Alergenele recombinate – metode de producere şi utilitatea în diagnosticul molecular şi în imunoterapia alergenică
Recombinant Allergens – Production Methods and Use in Molecular Diagnosis and Allergen Immunotherapy
Abstract
Although allergen extracts show some disadvantages, they are widely used both for in vivo diagnosis and for allergen immunotherapy. One way to overcome allergen extracts’ limitations is to use recombinant allergens. Selection of an appropriate expression system is dependent on the characteristics and intended application of the recombinant protein. The prokaryotic expression system is suitable for allergens that do not require post-translational modifications; however, there are allergens that need to be expressed in eukaryotic cells in order to acquire their biological activity. The use of these recombinant proteins ensures an increased specificity and sensitivity of the diagnosis and safer and more effective immunotherapy.Keywords
recombinant allergensprokaryotic expression systemeukaryotic expression systemmolecular diagnosismolecular allergen immunotherapyRezumat
Extractele alergenice sunt folosite pe scară largă atât în diagnosticul in vivo, cât şi în terapia bolilor alergice, deşi prezintă multiple dezavantaje. O soluţie pentru a evita limitările extractelor este utilizarea de alergene recombinate. Selecţia unui sistem de expresie proteică depinde de caracteristicile şi aplicaţiile proteinei recombinate. Alergenele care nu necesită modificări posttranslaţionale pot fi obţinute într-un sistem de expresie de tip procariot, dar există şi alergene care trebuie exprimate în celule eucariote pentru a-şi putea dobândi activitatea biologică. Utilizarea clinică a acestor proteine recombinate duce la creşterea specificităţii şi sensibilităţii diagnosticului şi asigură o imunoterapie mai sigură şi mai eficientă.Cuvinte Cheie
alergene recombinatesistem de expresie proteică de tip procariotsistem de expresie proteică de tip eucariotdiagnostic molecularimunoterapia alergenică molecularăIntroducere
În prezent, diagnosticul in vivo al alergiilor şi imunoterapia alergenică se bazează în mare pe extracte alergenice obţinute din surse naturale. Deşi calitatea acestor extracte a crescut în ultimele decenii prin standardizarea procedurii de obţinere, faptul că provin din surse naturale le face foarte eterogene, existând variaţii în ceea ce priveşte compoziţia şi cantitatea de molecule alergenice(1,2). De asemenea, extractele pot fi contaminate cu alergene provenite din alte surse sau pot conţine molecule bioactive care pot afecta stabilitatea acestora (enzime proteolitice care pot degrada alergenele)(3,4).
Tehnologiile de obţinere a alergenelor recombinate pot fi folosite pentru a înlătura neajunsurile extractelor alergenice. În ultimele decenii a fost produsă o mare varietate de alergene recombinate similare ca structură, funcţie şi răspuns imun cu omoloagele lor naturale, provenite de la plante, mucegaiuri, acarieni, insecte şi mamifere, folosind atât sisteme de expresie de tip procariot, cât şi de tip eucariot(5,6). Utilizarea clinică a acestor produse ajută la creşterea specificităţii şi sensibilităţii diagnosticului(7) şi asigură o imunoterapie mai sigură şi mai eficientă(8).
Primul pas în producerea alergenelor recombinate îl reprezintă obţinerea secvenţei de nucleotide care codifică proteina de interes. Folosind ARN mesager (ARNm) izolat din sursa alergenică, se sintetizează prin reacţia de polimerizare în lanţ în timp real (RT-PCR)(9,10) ADN complementar (ADNc), care mai apoi este clonat într-un vector specific sistemului de expresie care urmează să fie folosit.
În funcţie de complexitatea structurală şi funcţională a alergenelor, se pot folosi două sisteme de expresie, de tip bacterian şi de tip eucariot. Sistemul de tip bacterian este utilizat cu succes şi costuri reduse în cazul proteinelor cu structură simplă, care nu necesită modificări posttranslaţionale. Alergenele mai complexe din punct de vedere structural, cu glicozilări şi punţi disulfidice, sunt produse în sisteme de expresie de tip eucariot. Un avantaj al acestui sistem este purificarea simplă, redusă la o singură procedură, cromatografia de afinitate(1).
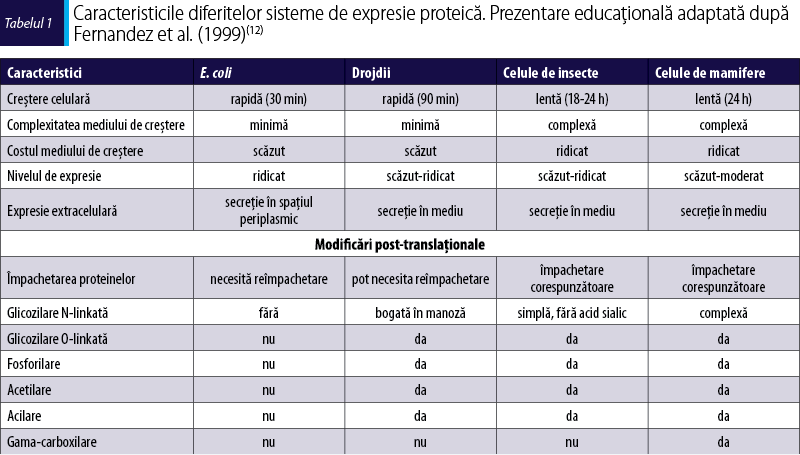
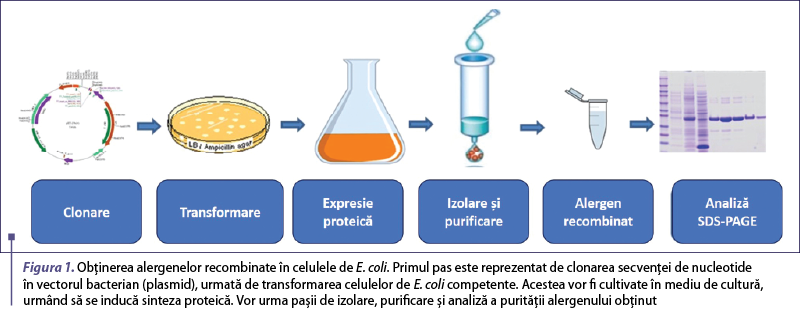
Acest articol analizează şi compară diferite sisteme de expresie folosite pentru producerea alergenelor recombinate (tabelul 1), precum şi utilizarea acestora în vederea îmbunătăţirii diagnosticului şi terapiei în bolile alergice.
Sistemul de expresie proteică de tip bacterian
Escherichia coli este una dintre cele mai utilizate gazde bacteriene pentru producerea cu randament ridicat de proteine recombinate. E. coli este un sistem convenabil şi versatil de obţinere a unei cantităţi mari de alergen recombinat, datorită ratei rapide de creştere într-un mediu de cultură ieftin şi uşor de preparat (figura 1). Cu toate acestea, este un sistem procariot căruia îi lipseşte mecanismul celular responsabil de procesele şi modificările posttranslaţionale, dezavantajul major fiind reprezentat de eşecul unor proteine recombinate de a dobândi structura secundară nativă, activă biologic. Adesea, supraproducţia de proteine străine în citoplasma E. coli determină o agregare a acestora, formând incluziuni celulare insolubile, care necesită solubilizare cu agenţi chatropici puternici, urmată de o reîmpachetare graduală pentru a deveni active funcţional(13). Un astfel de exemplu îl reprezintă alergenul major din chiparos, Cup a 1.02, care a fost exprimat în E. coli şi a necesitat solubilizare şi reîmpachetare pentru a redobândi forma reactivă IgE a alergenului(13).
Strategiile pentru creşterea calităţii şi randamentului de producere a proteinelor recombinate în celule bacteriene s-au concentrat pe îmbunătăţiri genetice, precum construirea unor vectori de expresie mai buni şi prin eliminarea genelor codificatoare de proteaze din tulpinile bacteriene. De asemenea, au fost create tulpini bacteriene capabile să faciliteze formarea punţilor disulfidice. Alergenul Ara h 2, din alune, a fost produs prin expresia într-o tulpină de E. coli modificată, Origami (DE3), care conţine în citoplasmă elemente oxidative care permit formarea punţilor disulfidice(14).

Sistemul de expresie procariot continuă să fie prima alegere când vine vorba despre producerea de alergene recombinate, numeroase alergene fiind produse în acest sistem (de exemplu, Ara h 1(15), Bet v 1(16), Fel d 1(17), Cor a 1(18)).
Sistemul de expresie proteică de tip eucariot
Avantajul major al sistemelor de expresie de tip eucariot este reprezentat de capacitatea de a realiza modificări posttranslaţionale, precum formarea legăturilor disulfidice, adăugarea de lipide şi carbohidraţi. Cele mai utilizate sisteme de expresie de tip eucariot sunt drojdiile, celulele de plante, celulele de insecte şi celulele de mamifere.
Având o rată de înmulţire ridicată şi necesităţi minime, sistemul de expresie în drojdii s-a dovedit util pentru producerea de alergene recombinate. Popularitatea crescută a acestui sistem se datorează procesului simplu şi randamentului ridicat de obţinere a proteinelor, precum şi capacităţii de a efectua modificări posttranslaţionale (formarea de legături disulfidice şi glicozilări). Organismul cel mai utilizat este Pichia pastoris, primele alergene exprimate fiind Cyn d 1(19), Alt a 1(20), Mus m 1(21) şi Bla g 4(22). Principalul dezavantaj al sistemului de expresie în drojdii este hiperglicozilarea proteinelor(23), modelele de glicozilare N- sau O-linkată putând fi diferite de cele ale proteinei native(24).
Sistemul de expresie în celule de insecte, asociat cu un vector baculovirus, devine tot mai des utilizat în domeniul producerii de alergene recombinate. Acest sistem se bazează pe capacitatea baculovirusului de a infecta celulele de insecte, având o specificitate ridicată în ceea ce priveşte tipul de gazdă infectată. Procesul de obţinere a proteinelor recombinate presupune clonarea genei de interes într-un vector bacterian (plasmid helper), cu care sunt mai apoi transformate celulele de E. coli competente care conţin bacmidul (plasmid care conţine baculovirusul fără gena de interes). Secvenţa de nucleotide care codifică alergenul de interes este apoi inserată în bacmid prin transpoziţie, rezultând un bacmid recombinat care va fi utilizat în transfecţia celulelor de insecte (figura 2) care vor asambla baculovirusul, continuându-se infecţia. Alergenul recombinat este secretat în mediul de cultură, de unde urmează a fi izolat şi purificat după 36-96 de ore de la infecţie. Baculovirusul distruge celulele de insecte, astfel că nu se poate realiza o producţie continuă, neîntreruptă.
Printre alergenele obţinute folosind sistemul cu baculovirus se numără: Api m 2(26), Fel d 1(27), Mal f 1(28). Toate aceste alergene recombinate prezintă o capacitate de legare a IgE şi activitate biologică similare cu cele ale proteinelor native.
Deoarece necesită echipamente sofisticate şi presupune costuri ridicate de producţie, cu randament scăzut, sistemul de expresie în celule de mamifere este foarte rar folosit (Per a 1(29)). Pentru expresie se mai pot utiliza şi plantele, proteinele putându-se acumula în cantităţi ridicate în diferite organe ale acestora (s-au obţinut Bet v 1 recombinat imunoreactiv şi două alergene din latex – Hev b 1 şi Hev b 3 –, folosind plante de tutun(30)).
Indiferent de sistemul de expresie proteică folosit, alergenele recombinate obţinute urmează numeroşi paşi de purificare cromatografică şi, fiind apoi analizate din punctul de vedere al identităţii, calităţii, cantităţii şi stabilităţii, prin metode precum electroforeza, spectrometria de masă şi dicroism circular. De asemenea, vor fi comparate cu omoloagele naturale în ceea ce priveşte activitatea biologică (reactivitate IgE, teste de activare a bazofilelor).
În cadrul proiectului INSPIRED (Strategii inovative pentru prevenţia, diagnosticul şi terapia afecţiunilor respiratorii induse de polenul de ambrozie), echipa de cercetători de la centrul OncoGen, Timişoara, a reuşit să producă alergene recombinate din polenul de Ambrosia artemisiifolia. Pentru alergenele mai simple din punct de vedere conformaţional (Amb a 8, Amb a 9, Amb a 10) s-a folosit sistemul de expresie de tip procariot, iar pentru cele mai complexe (Amb a 1, Amb a 4, Amb a 6), sistemul de expresie în celule de insecte. De asemenea, pentru câteva alergene (Amb a 3, Amb a 5, Amb a 11, Amb a 12) s-a mers în paralel cu ambele sisteme, pentru a stabili care este cel mai potrivit pentru obţinerea unei reactivităţi IgE comparabile cu cea a omologului natural.
Utilizarea alergenelor recombinate în diagnosticul alergiilor
Deşi testele diagnostice care conţin alergene recombinate se utilizează tot mai mult, cele bazate pe extracte naturale continuă să fie utilizate pe scară largă(31). Compoziţia unui extract alergenic influenţează puternic rezultatele oricărui test bazat pe acel extract. Din extractele disponibile comercial pot să lipsească alergene importante sau compoziţia lor alergenică poate varia considerabil de la un lot la altul, ceea ce poate da rezultate fals negative în cazul anumitor pacienţi. Aceste variaţii de concentraţie şi conţinut alergenic au fost demonstrate la soluţiile de testare cutanată prick (skin prick test, SPT) pentru extractul de acarian Dermatophagoides pteronyssinus(32), mucegai Alternaria alternata(33) şi alun(34), precum şi pentru produsele de imunoterapie alergenică sublinguală (SLIT) pentru acarian(35) şi polen de mesteacăn(36).
Alergenele recombinate bine caracterizate pot fi folosite ca marker în diagnostic pentru a realiza profilul de sensibilizare al pacienţilor alergici şi pentru a selecta strategiile terapeutice potrivite. Reactivitatea IgE la alergenele cu o reacţie încrucişată extrem de ridicată permite identificarea pacienţilor cu sensibilizare largă sau care sunt expuşi riscului de a dezvolta simptome alergice la mai multe surse. Pe de altă parte, alergenele specifice anumitor surse alergenice pot fi folosite pentru a identifica pacienţii cu sensibilizare reală(37). Pentru îmbunătăţirea calităţii unor extracte alergenice din latex(38), alun(39) şi venin de viespe(40), utilizate în diagnostic, s-a folosit procesul de spiking (adică adăugarea de alergene recombinate). Mai mult decât atât, alergenele recombinate reprezintă o alternativă la alergenele naturale pentru diagnosticul alergiilor, când alergenul natural poate fi izolat doar în cantităţi mici din sursa iniţială(41).
Utilizarea alergenelor recombinate în imunoterapia alergenică moleculară
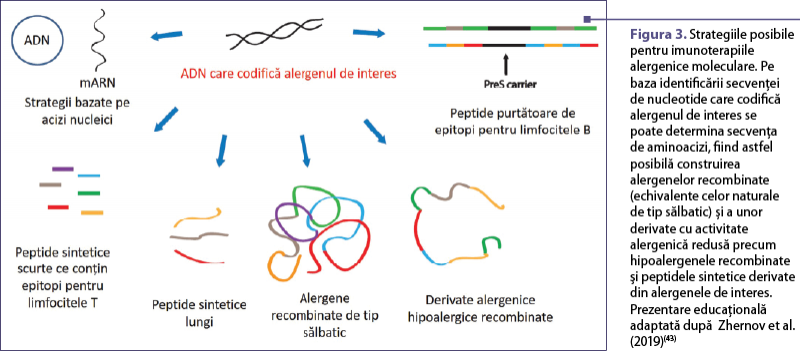
Izolarea primei secvenţe codificatoare a unui alergen a deschis drumul dezvoltării diferitelor tipuri de imunoterapii alergenice (AIT) moleculare(42) (figura 3). Printre acestea se numără alergenele recombinate de tip sălbatic (prezintă toate proprietăţile alergenelor naturale); peptidele sintetice care conţin epitopi alergen-specifici pentru limfocitele T (LT) (fără reactivitate IgE); acizi nucleici care codifică alergene; hipoalergene recombinate şi sintetice, cu alergenicitate scăzută şi capacitate redusă de legare IgE, dar cu epitopi alergen-specifici pentru LT(43).
Imunoterapiile personalizate bazate pe profilul de sensibilizare al fiecărui pacient sunt posibile, în teorie. Cu toate acestea, conform ghidurilor actuale, trebuie dovedită siguranţa şi eficienţa clinică pentru fiecare moleculă separat şi pentru posibilele combinaţii, ceea ce face această abordare imposibilă. De aceea imunoterapiile moleculare ar trebui să conţină un panel de alergene relevante clinic provenite de la sursa de interes.
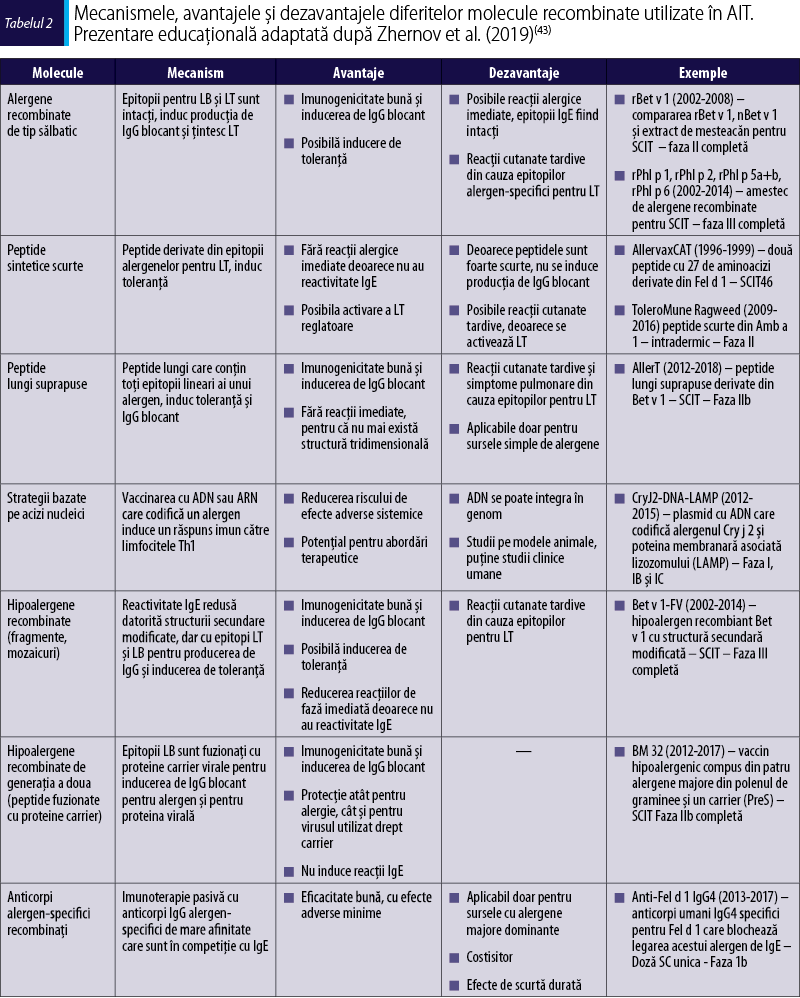
Avantajele şi dezavantajele diferitelor molecule recombinate folosite în AIT sunt prezentate în tabelul 2.
Concluzii
Pentru a determina sistemul optim de expresie a unui anumit alergen trebuie să se ţină cont de resursele disponibile, de timpul necesar, de calificarea personalului din laborator şi de natura alergenului. Există o experienţă vastă în ceea ce priveşte sistemul de expresie procariot, iar kiturile disponibile pe piaţă sunt mai variate, mai rapide şi mai puţin costisitoare decât cele pentru sistemele eucariote. Cu toate acestea, prezintă dezavantajul de a nu putea produce modificările posttranslaţionale necesare pentru proteinele complexe structural şi funcţional. Mai mult, aproape toate alergenele provin din surse eucariote şi este de preferat, în cazul în care activitatea biologică a alergenului recombinat depinde de conformaţia şi/sau de modificările posttranslaţionale, să se utilizeze sistemele eucariote de expresie proteică.
Tehnologia proteinelor recombinate permite atât producţia nelimitată a alergenului de interes, cât şi posibilitatea de a modifica structura şi proprietăţile intrinseci ale acestuia, devenind o alternativă din ce în ce mai atractivă pentru diagnosticul şi imunoterapia bolilor alergice.
Acest articol a fost elaborat în cadrul proiectului INSPIRED (Strategii inovative pentru prevenţia, diagnosticul şi terapia afecţiunilor respiratorii induse de polenul de ambrozia), cod SMIS 103662.
Bibliografie
- Chapman MD, Smith AM, Vailes LD, Arruda LK, Dhanaraj V, Pomés A. Recombinant allergens for diagnosis and therapy of allergic disease. J Allergy Clin Immunol. 2000;106(3):409-418.
- Valenta R, Kraft D. Recombinant allergen molecules: tools to study effector cell activation. Immunol Rev. 2001;179(1):119-127.
- van der Veen MJ, Mulderb M, Witteman AM, et al. False-positive skin prick test responses to commercially available dog dander extracts caused by contamination with house dust mite (Dermatophagoides pteronyssinus) allergens. J Allergy Clin Immunol. 1996;98(6):1028-1034.
- Nelson HS, Iklé D, Buchmeier A. Studies of allergen extract stability: the effects of dilution and mixing. J Allergy Clin Immunol. 1996;98(2):382-388.
- Scheiner O, Kraft D. Basic and practical aspects of recombinant allergens. Allergy. 1995;50(5):384-391.
- Schmid-Grendelmeier P, Crameri R. Recombinant allergens for skin testing. Int Arch Allergy Immunol. 2001;125(2):96-111.
- Bousquet J. In vivo and in vitro use of recombinant allergens. Allergy Clin Immunol News. 1994:54-59.
- Bousquet J, Lockey R, Malling HJ, et al. Special features – Allergen Immunotherapy: Therapeutic Vaccines for Allergic Diseases. Ann Allergy Asthma Immunol. 1998;81(5):401-405.
- Schmidt M, Zargari A, Holt P, et al. The complete cDNA sequence and expression of the first major allergenic protein of Malassezia furfur, Mal f 1. Eur J Biochem. 1997;246(1):181-185.
- Schmidt M, Walker RB, Hoffman DR, McConnell TJ. Nucleotide sequence of cDNA encoding the fire ant venom protein Sol i II. FEBS Lett. 1993;319(1-2):138-140.
- Schmidt M, Hoffman DR. Expression systems for production of recombinant allergens. Int Arch Allergy Immunol. 2002;128(4):264-270.
- Fernandez JM, Hoeffler JP. Gene Expression Systems: Using Nature for the Art of Expression. Elsevier; 1998.
- Rea G, Iacovacci P, Ferrante P, et al. Refolding of the Cupressus arizonica major pollen allergen Cup a1.02 overexpressed in Escherichia coli. Protein Expr Purif. 2004;37(2):419-425. doi:https://doi.org/10.1016/j.pep.2004.06.034
- Lehmann K, Hoffmann S, Neudecker P, Suhr M, Becker W-M, Rösch P. High-yield expression in Escherichia coli, purification, and characterization of properly folded major peanut allergen Ara h 2. Protein Expr Purif. 2003;31(2):250-259.
- Kleber-Janke T, Becker W-M. Use of modified BL21 (DE3) Escherichia coli cells for high-level expression of recombinant peanut allergens affected by poor codon usage. Protein Expr Purif. 2000;19(3):419-424.
- Swoboda I, Jilek A, Ferreira F, et al. Isoforms of Bet v 1, the major birch pollen allergen, analyzed by liquid chromatography, mass spectrometry, and cDNA cloning. J Biol Chem. 1995;270(6):2607-2613.
- van Ree R, van Leeuwen WA, Bulder I, Bond J, Aalberse RC. Purified natural and recombinant Fel d 1 and cat albumin in in vitro diagnostics for cat allergy. J Allergy Clin Immunol. 1999;104(6):1223-1230.
- Schenk S, Hoffmann‐Sommergruber K, Breiteneder H, et al. Four recombinant isoforms of Cor a 1, the major allergen of hazel pollen, show different reactivities with allergen‐specific T‐lymphocyte clones. Eur J Biochem. 1994;224(2):717-722.
- Smith PM, Suphioglu C, Griffith IJ, Theriault K, Knox RB, Singh MB. Cloning and expression in yeast Pichia pastoris of a biologically active form of Cyn d 1, the major allergen of Bermuda grass pollen. J Allergy Clin Immunol. 1996;98(2):331-343.
- De Vouge MW, Thaker AJ, Curran IH, et al. Isolation and expression of a cDNA clone encoding an Alternaria alternata Alt a 1 subunit. Int Arch Allergy Immunol. 1996;111(4):385-395. doi:10.1159/000237397
- Ferrari E, Lodi T, Sorbi RT, Tirindelli R, Cavaggioni A, Spisni A. Expression of a lipocalin in Pichia pastoris: secretion, purification and binding activity of a recombinant mouse major urinary protein. FEBS Lett. 1997;401(1):73-77.
- Vailes LD, Kinter MT, Arruda LK, Chapman MD. High-level expression of cockroach allergen, Bla g 4, in Pichia pastoris. J Allergy Clin Immunol. 1998;101(2):274-280.
- Bobrowicz P, Davidson RC, Li H, et al. Engineering of an artificial glycosylation pathway blocked in core oligosaccharide assembly in the yeast Pichia pastoris: production of complex humanized glycoproteins with terminal galactose. Glycobiology. 2004;14(9):757-766.
- Cereghino JL, Cregg JM. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol Rev. 2000;24(1):45-66.
- https://biotop.boku.ac.at/Projects/2015/Grabherr2015c.html.
- Soldatova LN, Crameri R, Gmachl M, et al. Superior biologic activity of the recombinant bee venom allergen hyaluronidase expressed in baculovirus-infected insect cells as compared with Escherichia coli. J Allergy Clin Immunol. 1998;101(5):691-698.
- Guyre P, Goldstein J, Wu Z, Sun A. Recombinant cat allergen, Fel dI, expressed in baculovirus for diagnosis and treatment of cat allergy. November 2002.
- Zargari A, Schmidt M, Lundberg M, Scheynius A, Whitley P. Immunologic characterization of natural and recombinant Mal f 1 yeast allergen. J Allergy Clin Immunol. 1999;103(5):877-884.
- Wu CH, Lee MF, Wang NM. Expression of the American cockroach Per a 1 allergen in mammalian cells. Allergy. 2000;55(12):1179-1183.
- Breiteneder H, Krebitz M, Wiedermann U, et al. Rapid production of recombinant allergens in Nicotiana benthamiana and their impact on diagnosis and therapy. Int Arch Allergy Immunol. 2001;124(1-3):48-50.
- Tscheppe A, Breiteneder H. Recombinant Allergens in Structural Biology, Diagnosis, and Immunotherapy. Int Arch Allergy Immunol. 2017;172(4):187-202. doi:10.1159/000464104
- Casset A, Mari A, Purohit A, et al. Varying allergen composition and content affects the in vivo allergenic activity of commercial Dermatophagoides pteronyssinus extracts. Int Arch Allergy Immunol. 2012;159(3):253-262.
- Kespohl S, Maryska S, Zahradnik E, Sander I, Brüning T, Raulf‐Heimsoth M. Biochemical and immunological analysis of mould skin prick test solution: current status of standardization. Clin Exp Allergy. 2013;43(11):1286-1296.
- Akkerdaas JH, Wensing M, Knulst AC, Aalberse RC, Hefle SL. In vitro and in vivo characterization of hazelnut skin prick test extracts. Arb aus dem Paul-Ehrlich-Institut (Bundesamt fur Sera und Impfstoffe) zu Frankfurt aM. 2003;(94):87-95.
- van Ree R. Indoor allergens: relevance of major allergen measurements and standardization. J Allergy Clin Immunol. 2007;119(2):270-277.
- Moreno Benítez F, Espinazo Romeu M, Letrán Camacho A, Mas S, García‐Cózar FJ, Tabar AI. Variation in allergen content in sublingual allergen immunotherapy with house dust mites. Allergy. 2015;70(11):1413-1420.
- Kazemi-Shirazi L, Niederberger V, Linhart B, Lidholm J, Kraft D, Valenta R. Recombinant Marker Allergens: Diagnostic Gatekeepers for the Treatment of Allergy. Int Arch Allergy Immunol. 2002;127(4):259-268. doi:10.1159/000057742
- Lundberg M, Chen Z, Rihs H, Wrangsjö K. Recombinant spiked allergen extract. Allergy. 2001;56(8):794-795.
- Andersson K, Ballmer‐Weber BK, Cistero‐Bahima A, et al. Enhancement of hazelnut extract for IgE testing by recombinant allergen spiking. Allergy. 2007;62(8):897-904.
- Yoshida N, Hirata H, Watanabe M, et al. Improved sensitivity to venom specific-immunoglobulin E by spiking with the allergen component in Japanese patients suspected of Hymenoptera venom allergy. Allergol Int. 2015;64(3):248-252.
- Mas S, Boissy P, Monsalve RI, et al. A recombinant Sal k 1 isoform as an alternative to the polymorphic allergen from Salsola kali pollen for allergy diagnosis. Int Arch Allergy Immunol. 2015;167(2):83-93.
- Valenta R, Ferreira F, Focke-Tejkl M, et al. From allergen genes to allergy vaccines. Annu Rev Immunol. 2009;28:211-241.
- Zhernov Y, Curin M, Khaitov M, Karaulov A, Valenta R. Recombinant allergens for immunotherapy: state of the art. Curr Opin Allergy Clin Immunol. 2019;19(4):402-414.

Sindrom de alergie orală în rinita alergică la polen de platan
Carmen Saviana Ganea, Mariana Vieru, Florin-Dan Popescu
Reactivitatea încrucişată dintre polenul de Platanus şi alimente de origine vegetală este descrisă în literatura de specialitate, dar componentele alergenice cu reactivitate încrucişată/cross-reactive nu au fost stabilit...
Componente alergenice din alimente declarabile ca surse de alergene în UE, disponibile în metodele de imunoanaliză multiplex
Florica Popescu, Mariana Vieru, Florin-Dan Popescu, Carmen Bunu-Panaitescu
Diagnosticul alergologic molecular în alergiile alimentare este o abordare modernă, utilizată pentru a caracteriza componentele moleculare ale alergenelor alimentare din punctul de vedere al răspunsur...
Inhibitorii de N-glicani în metodele de imunoanaliză pentru IgE alergen-specifice serice
Florin-Dan Popescu, Mariana Vieru, Carmen Saviana Ganea, Carmen Bunu-Panaitescu
Determinanţii carbohidraţi cu reactivitate încrucişată/cross-reactivi (CCD) reprezintă N-glicani fucozilaţi/xilozilaţi prezenţi în polenuri şi alimente de origine vegetală, latex şi veninuri de himenoptere. Anticorpii Ig...
Implementarea unei soluţii pentru îmbunătăţirea calităţii vieţii pacienţilor alergici la bioparticule
Imbrea Florin, Chiţu Cătălin , Săndesc Dorel , Laura Haidar, Maria Roxana Buzan, Carmen Panaitescu
Articolul se concentrează pe un proiect inovator desfăşurat în Timişoara, care urmăreşte identificarea, monitorizarea şi analiza polenurilor şi sporilor de mucegaiuri prezente în aer....
Relevanţa clinică a profilinelor în bolile alergice
Maria Roxana Buzan, Manuela Grijincu, Lauriana-Eunice Zbîrcea, Laura Haidar, Tudor Paul Tamaş, Monica Cotarcă, Carmen Panaitescu
Profilinele sunt proteine care leagă actina, având rol în organizarea citoscheletului. Acestea sunt prezente în citoplasma tuturor celulelor eucariote, atât din surse animale, cât şi din plante. Cu to...