Din aceeași categorie

Introducere
Fungusul Alternaria alternata a devenit prima dată cunoscut comunităţii ştiinţifice sub numele de Alternaria tenuis. În 1817, Christian Gottfried Daniel Nees von Esenbeck a stabilit genul Alternaria şi a descris Alternaria tenuis ca specie caracteristică. În 1912, micologul austriac Karl von Keißler a redenumit Alternaria tenuis ca Alternaria alternata. Abia în 1930 s-a stabilit că sporii de Alternaria provoacă astm alergic. Acest mucegai este considerat în prezent sursă importantă de aeroalergene fungice sensibilizante(1,2), cu impact uneori subestimat în alergiile respiratorii. Alternaria alternata este cea mai frecventă şi abundentă specie de fungi care provoacă astm alergic, în special la copii, şi este asociată cu severitatea şi persistenţa simptomelor respiratorii(3,4,5).
Prevalenţa alergiei mediate IgE la fungi în populaţia generală este de aproximativ 3-10%(6), iar sensibilizarea la Alternaria alternata la atopici variază între 3% şi 40%(7), în funcţie de regiunea climatică şi populaţia testată.
Alternaria alternata face parte din familia Pleosporaceae, ordinul Pleosporales, clasa Dothideomycetes a încrengăturii Ascomycota. Este în principal un fungus de exterior ai cărui spori diseminează în aerul cald şi uscat, astfel că în regiunile temperate concentraţia acestora atinge valori maxime la sfârşitul verii şi începutul toamnei. Alternaria se găseşte de asemenea în medii de interior umede, în case insuficient ventilate(1,5,8). Sensibilizarea la Alternaria este asociată cu un risc mai mare de exacerbări ale astmului în perioada mai-august în sudul Europei şi în iulie-august în nordul continentului(9,10).
Sporii de Alternaria alternata sunt esenţiali pentru supravieţuirea şi dispersia fungică şi sunt rezistenţi la condiţii climatice nefavorabile(11). Alergenicitatea semnificativă a sporilor de Alternaria este relevată de faptul că pragul de concentraţie a sporilor din aer necesar pentru a provoca simptome mediate IgE a fost estimat la 100 spori/m3, comparativ cu 3000 spori/m3 pentru Cladosporium(12). Numărul de spori de Alternaria din aer este adesea de 1000 de ori mai mare decât cel al grăuncioarelor de polen, iar expunerea este adesea mai lungă ca durată. Există o corelaţie clară între concentraţiile atmosferice ridicate ale anumitor spori de mucegai în timpul furtunilor electrice şi o creştere a internărilor în spital din cauza exacerbărilor astmului(13). Mai mult, concentraţiile crescute de spori de Alternaria (mai mari de 1000 spori/m3 aer) se pot asocia cu creşterea riscului de deces datorat astmului în mediul urban(14). În plus, fungi precum Alternaria activează şi sistemul imunitar înnăscut şi pot amplifica inflamaţia provocată de aeroalergene neînrudite, precum polenurile de graminee sau iarba pârloagelor(13,15).
Alternaria reprezintă o sursă importantă de molecule alergenice fungice, implicând diferite structuri moleculare conservate, inclusiv enzime, componente ale peretelui celular sau proteine cu reactivitate încrucişată(16). Deşi sunt descrise sute de specii care aparţin genului Alternaria, doar două dintre ele au implicaţii alergologice semnificative, şi anume Alternaria alternata şi Ulocladium chartarum (Alternaria chartarum)(17). Extractele naturale de Alternaria alternata prezintă variaţii mari în ceea ce priveşte potenţa alergenică(18).
Alergenele moleculare ale fungusului Alternaria alternata
Identificarea şi caracterizarea tuturor moleculelor alergenice din Alternaria alternata au o importanţă clinică particulară(12). Extractele alergenice din sporii şi hifele de Alternaria sunt încă utilizate în practică, dar au compoziţie moleculară variabilă şi sunt insuficient standardizate. Înainte de 1990, datele ştiinţifice referitoare la alergenele relevante din Alternaria alternata erau foarte limitate. Între timp, până în 2008, au fost identificate 13 componente moleculare. Nouă dintre acestea – şi anume, enolaza, manitol-dehidrogenaza, aldehid-dehidrogenaza, flavodoxina, proteinele P1 şi P2 ribozomale, proteina de şoc termic, factorul de transport nuclear şi glutation-transferaza – au fost identificate în Alternaria alternata, dar şi în fungusul înrudit Cladosporium herbarum(7). La mijlocul anilor 1990 au fost obţinute o serie de alergene proteice fungice recombinante şi au fost deduse o parte din funcţiile lor biologice(19,20). Alt a 1 a fost identificat ca alergen major al Alternaria alternata specific pentru Pleosporaceae. În soluţiile comerciale pentru testare cutanată prick s-au depistat niveluri diferite de Alt a 1, iar alergenele cross-reactive au început să fie caracterizate. Până în 2014, au fost izolate 16 alergene din Alternaria alternata, dar funcţiile biologice ale Alt a 1 şi ale câtorva alergene minore au rămas incomplet elucidate sau necunoscute(1).
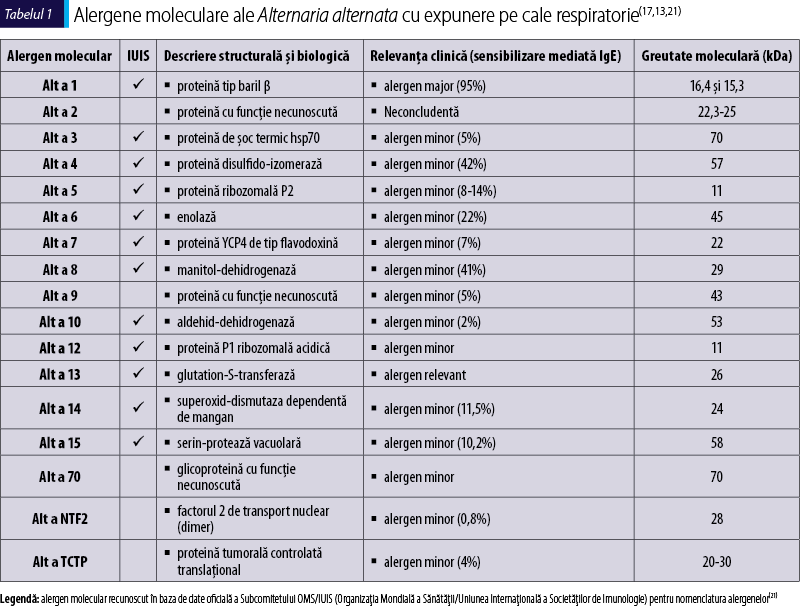
Actualmente, alergenele moleculare identificate în Alternaria alternata sunt în număr de 17: Alt a 1 (alergen major), Alt a 2 (neconcludent) şi alergene minore (Alt a 3, Alt a 4, Alt a 5, Alt a 6, Alt a 7, Alt a 8, Alt a 9, Alt a 10, Alt a 12, Alt a 13, Alt a 14, Alt a 15, Alt a NFT2, Alt a TCTP, Alt a 70 kDa)(13).
Sporii şi fragmentele fungice sunt componente obişnuite ale aerului din medii de exterior şi interior care pot produce alergii respiratorii(22). Faptul că extractele naturale din spori şi micelii sunt capabile să inducă simptome alergice la teste de provocare nazală oferă dovada că alergene fungice relevante sunt prezente în ambele forme de existenţă(23).
Alt a 1, alergenul major al Alternaria alternata, este o proteină relativ mică (157 de aminoacizi), detectată mai ales în spori, din care este eliberată la începutul germinării(24).
Prezenţa Alt a 1 în spori şi micelii a fost demonstrată prin microscopie electronică cu transmisie şi imunodetecţie cu aur coloidal(22,25). În spori a fost depistat în peretele celular, iar la acest nivel, în stratul de melanină al sporilor mai maturi, dar nesemnificativ în citoplasmă, în hife sau în sporii mai tineri(11). Localizarea distinctă a Alt a 1 în peretele celular este relevată când mucegaiul este crescut în condiţii care le mimează pe cele naturale cu expunere în aer. În fungii dezvoltaţi în medii de cultură lichide artificiale, Alt a 1 are localizare difuză în citoplasmă şi în peretele celular, posibil datorită dislocării lui în mediu lichid, fenomen care s-a constatat că provoacă difuzia alergenelor şi din grăuncioarele de polen prin hidratare(15).
Concentraţiile din aer ale Alt a 1 şi ale sporilor de Alternaria se corelează semnificativ, prin urmare se poate presupune că majoritatea componentei alergenice Alt a 1 din atmosferă este derivată din sporii fungici intacţi. Deoarece sporii contribuie la infectarea plantelor, iar Alt a 1 este o proteină implicată în patogeneza plantelor necesară pentru a bloca apărarea acestora şi pentru a favoriza contaminarea lor fungică, nu este surprinzător faptul că Alt a 1 este prezent în cele mai mari concentraţii în pereţii celulari ai sporilor maturi şi germinanţi(26).
Alt a 1 este bine caracterizat ca alergen major şi biomarker al sensibilizării primare mediate IgE la Alternaria alternata. rAlt a 1 este alergenul molecular recombinant utilizat în mod obişnuit pentru diagnosticul de sensibilizare la Alternaria. Anticorpii IgE specifici faţă de Alt a 1 sunt depistaţi la peste 90% până la 98% dintre pacienţii sensibilizaţi la acest fungus(5,27,28,29,30). Este un biomarker de sensibilizare puternic asociat cu astmul persistent, dar şi un factor de risc pentru exacerbări şi astm alergic sever(13,24). Sensibilizarea la Alt a 1 poate fi depistată la un sfert dintre pacienţii cu dermatită atopică, în special forme moderate şi cu rinită sau astm alergic coexistent(16).
Alt a 1 din Alternaria alternata este un alergen cu structură aparte şi activităţi biologice neelucidate, inclus în propria familie de proteine. Studiile realizate cu alergenul omolog Alt b 1 din fungusul necrotrofic Alternaria brassicicola au relevat faptul că se exprimă preferenţial în sporii care germinează pe frunze de Arabidopsis, dar nu şi în sporii care germinează în apă, susţinând rolul în patogeneza fungică a plantelor, iar capacitatea de a inhiba proteinele vegetale PR5 de tip taumatină sugerează roluri biologice în dezvoltarea interacţiunii plante-fungi. În plus, se consideră că ar avea implicaţii în germinarea sporilor(18,31,32,33,34,35,36).
Alt a 1 este o proteină unică de tip baril beta, formată din 11 lanţuri beta cu conformaţie spaţială asemănătoare unui butoi. Alt a 1 este un alergen specific, fără reactivitate încrucişată cu alte alergene fungice care au alte structuri proteice. Câteva alergene cu omologie Alt a 1 au fost depistate exclusiv la fungi din familia Pleosporaceae(10,13,24,33,34,36). Dintre moleculele alergenice cu omologie Alt a 1 menţionăm: Ulo c 1 din Ulocladium chartarum(17), Ulo b 1 din Ulocladium botrytis şi Ste b 1 din Stemphylium botryosum(37). Alt a 1 este un biomarker pentru sensibilizarea la fungi din familia Pleosporaceae a clasei Dothideomycetes, neincluzând Curvularia spp(34,38). Expresia eficientă a genei care codifică Alt a 1 a fost evidenţiată în Alternaria alternata şi în alte genuri importante din familia Pleosporaceae, cum ar fi Stemphylium botryosum şi Ulocladium chartarum(39).
Structura Alt a 1 este a unui dimer termostabil de 30 kDa, care se disociază în subunităţi de 16,4 şi 15,3 kDa în condiţii reducătoare sau de pH acid. Dimerul cu formă de „fluture” este stabilizat printr-o punte disulfidică între Cys30 din ambii monomeri şi prin interacţiuni hidrofobe şi polare(13,24,36,40).
Dimerul stabil prezintă epitopi cu o orientare adecvată pentru cross-linkarea IgE. Deşi Alt a 1 are o structură dimerică, ipoteza că dimerizarea este o condiţie prealabilă importantă pentru alergenicitate nu este susţinută(12,41). Epitopii IgE liniari K41-P50 şi Y54-K63 identificaţi în Alt a 1 sunt localizaţi pe dimer cu expunere care permite interacţiunea cu IgE. În schimb, oligomerizarea tetramerică a Alt a 1, stabilizată de ligandul său nativ flavonol metilat, asemănător flavonolului vegetal quercetină, ascunde epitopii de legare, blocând interacţiunea cu anticorpii IgE(13,24,42,43,44,45). Alt a 1 nu este doar alergen major din Alternaria alternata, dar conţine şi cei mai importanţi epitopi IgE specifici ai acestui fungus(38).
Speciile de Alternaria nu sunt termotolerante şi nu pot germina în căile respiratorii, de aceea acţionează ca aeroalergene la care expunerea are variaţii sezoniere(46). Alergenul Alt a 1 din sporii inhalaţi este eliberat extrem de rapid şi în cantităţi importante la un pH fiziologic relevant în special în condiţii uşor acide. Alt a 1 induce producerea de IL-8, CXCL1, CXCL2, CXCL3 şi CCL2/MCP-1 în celulele epiteliale ale căilor respiratorii într-o manieră dependentă de receptorii de recunoaştere toll-like TLR2 şi TLR4 şi proteinele adaptoare MyD88 şi TIRAP, ceea ce sugerează că Alt a 1 ar putea fi recunoscut de macrofagele alveolare(44). Alt a 1 în forma sa tetramerică şi în prezenţa ligandului interacţionează şi cu receptorul de suprafaţă SLC22A17 pentru siderocalină la nivelul celulelor epiteliale bronşice, suferă endocitoză mediată de receptor şi, fără modificări evidente ale integrităţii epiteliale, induce expresia de alarmine IL-33 şi IL-25(45). Alarminele promovează recrutarea şi activarea celulelor limfoide înnăscute de tip 2 (ILC2), care au un rol important în dezvoltarea răspunsurilor de tip 2 induse de Alternaria(13).
Expunerea pacienţilor la alergenul major Alt a 1 are loc în principal prin expunerea la spori de Alternaria alternata în mediul exterior la sfârşitul verii şi toamna(6). Sporii de Alternaria sunt principala sursă de Alt a 1 în aerul liber. Ei variază ca mărime de la aproximativ 20 µm la 200 µm şi sunt de multe ori mai mari decât fragmentele fungice de dimensiuni micronice. Doar aproximativ 25% dintre fragmentele de hife prezintă alergene detectabile, iar concentraţiile lor aeropurtate le depăşesc pe cele ale sporilor de numai aproximativ două-trei ori. Impactul particulelor fungice mai mici decât sporii (fragmente de hife, conidiofori, spori fragmentaţi) nu poate fi neglijat totuşi, deoarece acestea pot creşte concentraţia atmosferică totală de Alt a 1 şi, datorită dimensiunilor lor mici, pot rămâne în aer mai mult timp şi penetrează mai profund şi mai uşor în tractul respirator inferior. Există o variabilitate a cantităţii de Alt a 1 detectată per spor, proporţia de Alt a 1 în sporii din fracţiunea PM2.5-10 fiind cu aproximativ 30% mai mare decât cea din PM>10, probabil din cauza fragmentelor de hife. De aceea s-a recomandat recent includerea metodelor de măsurare a alergenelor moleculare în monitorizarea aerobiologică. Cele mai mari niveluri de Alt a 1 în aer sunt înregistrate în timpul zilelor însorite, calde şi uscate, când condiţiile meteorologice favorizează mişcarea ascendentă a curenţilor de aer (temperatură ridicată, durată mai mare a perioadelor însorite, prezenţa norilor cumulus, punctul de rouă şi deficitul presiunii vaporilor), cunoscute a influenţa în mod pozitiv concentraţiile sporilor de Alternaria. Fragmentarea sporilor de Alternaria prin leziuni mecanice în timpul recoltării automatizate a cerealelor sau în cazul furtunilor cu curenţi de aer la nivelul ierburilor pare a fi mai importantă decât ruptura lor prin şoc osmotic(26).
Alergenele din grupul 1 al β-expansinelor prezente în grăuncioarele de polen de graminee şi în paie sunt capabile de legare fizică la sporii de Alternaria care conţin Alt a 1. De aceea, sporii de mucegai prezenţi pe iarbă pot acţiona ca transportatori de alergene polinice aeropurtate. Exacerbările astmului din timpul toamnei la pacienţii alergici la polen de Poaceae pot fi provocate prin inhalarea de spori fungici care transportă alergene de graminee din Grupul 1. Un astfel de amestec este favorizat în timpul furtunilor electrice, deoarece acestea pot creşte semnificativ cantitatea de particule de polen paucimicronice şi de spori fungici în aerul respirabil. În plus, gradul ridicat de cosensibilizare la alergenele din polenul de graminee şi cele din Alternaria contribuie la agravarea simptomatologiei(47).
Alt a 1 este şi un biomarker al condiţiilor optime de cultură pentru producerea extractelor naturale de Alternaria alternata destinate diagnosticului alergologic şi imunoterapiei specifice. Expresia Alt a 1 în timpul creşterii în condiţii de cultură poate fi evaluată prin metode de imunoanaliză enzimatică de tip sandwich în fază solidă şi imunoblot(22).
În plus, Alt a 1 este şi un biomarker al contaminării fructelor şi legumelor. Determinarea Alt a 1 se poate realiza prin metoda ELISA sau PCR. Alternaria solani şi Alternaria alternata pot infecta tomatele, cartofii sau citricele, producând alternarioza cu pete brune-negre(48,49). Este posibil ca sporii de Alternaria să fie prezenţi în fructe/legume fără a dezvolta hife, iar sporii să producă componente moleculare implicate în alergenicitate, cum ar fi Alt a 1. Acest alergen al fungului saprotrofic poate fi detectat în fructe, în ciuda faptului că dezvoltarea hifelor nu este observată după mai multe zile postinoculare. Alt a 1 interacţionează cu proteina PR5 de tip taumatină din fructul kiwi, iar ambele proteine se colocalizează în pulpă, interacţiunea ducând la inhibarea activităţii enzimatice a proteinei PR5. Acest efect nu se limitează la kiwi, ci apare şi la PR5 din alte fructe, cum ar fi piersica sau banana. Fenomenul de cosensibilizare la Alt a 1 şi Act d 2 (alergenul major de tip taumatină din fructul kiwi) poate fi determinat de consumul de fructe contaminate cu Alternaria, dar aparent optime din punct de vedere organoleptic. Alimentele infectate fungic pot fi şi cauza unor reacţii alergice(50).
În cazul sindromului Alternaria-spanac a fost incriminat un alergen comun fungic şi vegetal de 30 kDa(51), suspicionat a fi alergenul molecular Alt a 1(52).
Nivelurile de IgE specifice raportate faţă de componenta nativă nAlt a 1 sau recombinantă rAlt a 1 au arătat o corelaţie semnificativă, cu sensibilitate şi specificitate similare(53). Alt a 1 este considerat cel mai util alergen molecular specific pentru evaluarea sensibilizării mediate IgE la Alternaria(54), alături de enolaza cross-reactivă Alt a 6, importantă în contextul în care reactivitatea încrucişată este o componentă semnificativă a sensibilizării la fungi(55,56).
rAlt a 1 este singurul alergen molecular utilizat actualmente în practica alergologică atât în metodele in vitro de imunoanaliză singleplex, cât şi în cele multiplex pentru diagnosticul molecular al alergiei respiratorii la Alternaria alternata(57,58). În studii experimentale din trecut, alergenele moleculare fungice native şi recombinante, nAlt a 1 şi rAlt a 1, cu sau fără Alt a 6, au fost utilizate şi pentru diagnosticul fiabil in vivo al sensibilizării la fungusul Alternaria alternata prin testare cutanată prick(53,59). Extractele comerciale naturale de Alternaria pentru testare cutanată alergologică prick conţin concentraţii variabile de Alt a 1 (în funcţie de producător, de la 3,1 µg/mL la 441,1 µg/mL), iar unele alergene moleculare, precum Alt a 2, Alt a 3, Alt a 6 şi Alt a 8, pot să nu fie detectate în aceste soluţii pentru teste alergologice in vivo(18).
Componenta alergenică Alt a 1 poate fi discutată nu numai din perspectiva diagnosticului molecular de precizie, ci şi al imunoterapiei specifice în alergia respiratorie mediată IgE, inclusiv îmbunătăţirea acurateţei indicaţiei acesteia(5,16,56). Pe lângă datele ştiinţifice legate de imunoterapia cu extracte fungice naturale(60,61,62,63,64,65,66), există studii clinice de imunoterapie cu alergenul major Alt a 1 natural purificat(67,68). Imunoterapia cu rAlt a 1 poate reprezenta de asemenea una dintre strategiile terapeutice specifice de viitor(69). Utilizarea rAlt a 1 sau nAlt a 1 pentru imunoterapia alergenică elimină expunerea la potenţiale proteine complexe, precum micotoxine, alţi metaboliţi secundari şi fragmente de perete celular cu chitină, manani, β-1,3-glucani, care pot genera răspunsuri proinflamatoare(44, 70).
Alt a 2 în forma sa recombinantă a fost obţinut prin clonare moleculară şi a fost iniţial încadrat ca alergen major, considerându-se relevant la 61% dintre pacienţii sensibili la Alternaria(71), dar ulterior a fost reevaluat ca alergen minor, sensibilizarea mediată IgE la această componentă moleculară fungică nefiind detectată în unele studii. Este considerată proteină conservată în Alternaria cu secvenţă de 190 de aminoacizi şi greutate moleculară de 22,3-25 kDa(7,13,18,53,71,72,73). Această moleculă alergenică are funcţie biologică necunoscută, iar relevanţa clinică este neconcludentă(13).
Alt a 3 este o proteină de şoc termic menţionată ca hsp70, cu greutate moleculară de 70 kDa. IgE specifice serice faţă de componenta alergenică Alt a 3 sunt prezente la 5% dintre pacienţii sensibilizaţi la Alternaria alternata. Pentru acest fungus au fost clonate secvenţele hsp70 care codifică fragmentele de legare IgE cu omologie structurală importantă (72%) cu cele pentru alergenul Cla h hsp70 din Cladosporium herbarum(74). Alt a 3 nu este o proteină exprimată în mod constitutiv, ci un şaperon molecular care protejează celulele împotriva stresului oxidativ şi termic(75), care este exprimat în timpul sporulării fungice(76). Extractele din spori pot conţine niveluri mai mari de proteină hsp70 în comparaţie cu cele din micelii crescute în condiţii bogate din punct de vedere nutriţional. Alte exemple de alergene moleculare cu potenţial cross-reactiv induse de stres sunt: Pen c 19 (hsp 70) din Penicillium citrinum, Mala s 10 (hsp 70) din Malassezia sympodialis şi Asp f 12 (hsp P90) din Aspergillus fumigatus(77). Caracterul inductibil prin stres al acestui alergen subliniază nevoia de extracte fungice care să conţină un număr semnificativ de alergene obţinute în condiţii nutriţionale variate şi compararea expresiei hsp70 în extracte de spori şi micelii de Alternaria alternata(74).
Alt a 4 este o enzimă de 57 kDa, disulfid-izomeraza, care leagă IgE din serurile a 42% dintre pacienţii sensibilizaţi la Alternaria alternata(7,13,19,20,73). Proteina disulfid-isomerază (PDI) este o enzimă a reticulului endoplasmatic al eucariotelor şi a periplasmei procariotelor, care catalizează oxidarea, reducerea sau izomerizarea punţilor disulfurice între resturile de cisteină, cu facilitarea plierii proteinelor în structura tridimensională terţiară(78,79,80). Molecula de Alt a 4 conţine cel puţin un domeniu catalitic de tip tioredoxină ca situs activ(56).
Alt a 5 este o proteină P2 ribozomală acidică cu greutate moleculară de 11 kDa care leagă anticorpi IgE din serurile a 8-14% dintre pacienţii sensibilizaţi la Alternaria alternata(7). Alergenul este o ribonucleoproteină fosforilată cu o secvenţă completă de 113 aminoacizi. Regiunile C-terminale ale Alt a 5 prezintă probabil epitopi de legare a IgE(74). Proteina P2 ribozomală acidică 60S este un constituent structural al subunităţii mari a ribozomulului care are un rol important în etapa de elongare a sintezei proteinelor(81). Alt a 5 prezintă omologie extinsă cu Cla h 5, alergenul ribozomal proteic P2 al Cladosporium herbarum(19,82). Alte alergene fungice de tipul proteinei ribozomale P2 cu potenţial cross-reactiv sunt: Asp f 8 din Aspergillus fumigatus şi Fus c 1 din Fusarium culmorum(12,21). Este important de menţionat că sensibilizarea la alergene din mucegaiuri prin expunere la nivelul căilor respiratorii poate fi asociată ulterior cu anafilaxie după ingerarea orală de proteine fungice cu reactivitate încrucişată, aşa cum a fost cazul unui pacient astmatic cu reacţii severe de hipersensibilitate la micoproteina comestibilă comercială din Fusarium venenatum. Alergenul proteic P2 ribozomal al acestui fungus este implicat cu o mare probabilitate, din cauza potenţialului său de reactivitate încrucişată cu alergenul Fus c 1 din Fusarium culmorum(83). Mai mult, regiunea C-terminală a proteinelor ribozomale P2 prezintă omologie cu unele autoantigene implicate în lupusul eritematos sistemic(84). Au fost descrise răspunsuri IgE faţă de proteina ribozomală umană Hom s P2 în această afecţiune autoimună sistemică(85).
Alt a 6 (denumire anterioară abandonată: Alt a 11) este o enolază, cunoscută şi ca fosfopiruvat-hidratază sau 2-fosfo-D-glicerat-hidroliază, enzimă glicolitică care catalizează interconversia 2-fosfo-D-gliceratului (2PGA) în fosfoenolpiruvat (PEP)(1). Considerată iniţial enzimă citosolică implicată în glicoliză şi gluconeogeneză, enolaza este recunoscută actualmente ca proteină multifuncţională care poate apărea în diferite compartimente celulare(86,87). Enolaza este prezentă de la bacterii la mamifere, iar secvenţa sa este foarte bine conservată pe parcursul evoluţiei(88). Enzima nativă purificată Alt a 6 are greutate moleculară de 47 kDa, posedă termostabilitate ridicată şi leagă anticorpi IgE din serurile a 8% dintre subiecţii sensibilizaţi la fungus. De aceea, este considerat alergen minor, în contextul în care 22% dintre pacienţii alergici la Alternaria alternata reacţionează în mod specific cu alergenul recombinant de 45 kDa rAlt a 6(7,19,89,90,91).
rAlt a 6 este utilizat ca alergen cross-reactiv în metodele de imunoanaliză multiplex pentru evaluarea moleculară a alergiei respiratorii la Alternaria alternata(57,58). Sensibilizarea la Alt a 6 a fost observată şi la pacienţii cu dermatită atopică, mai frecvent în formele moderat-severe şi cu astm coexistent, nefiind depistată în formele uşoare de eczemă(16). Enolazele au fost identificate iniţial ca alergene în Saccharomyces cerevisiae(92) şi Candida albicans(93). În prezent, ele sunt considerate panalergene fungice cu reactivitate încrucişată semnificativă(7). Principalele enolaze fungice cross-reactive sunt: Alt a 6 din Alternaria alternata, Cla h 6 din Cladosporium herbarum, Asp f 22 din Aspergillus fumigatus, Pen c 22 din Penicillium citrinum, Cur l 2 din Curvularia lunata şi Rho m 1 din Rhodotorula mucilaginosa(12,21,91,94). Cross-reactivitatea dintre enolazele fungice (Alt a 6, Cla h 6) şi din latexul de Hevea brasiliensis (Hev b 9) este discutată în sindromul latex-mucegai(95).
Alt a 7 este un alergen minor de 22 kDa din Alternaria alternata, cu omologie structurală cu proteina YCP4 de tip flavodoxină din drojdia Saccharomyces cerevisiae. Alt a 7 se leagă in vitro la IgE serice de la 7% dintre persoanele sensibilizate la Alternaria alternata(7,19,73). În schimb, testarea cutanată in vivo cu Alt a 7 recombinant nu a produs rezultate pozitive la un lot redus de subiecţi alergici la Alternaria(59). Secvenţa completă de 204 aminoacizi a Alt a 7 este cunoscută, dar funcţia rămâne neidentificată(73). Proteina YCP4 acţionează ca reglator transcripţional(96) şi are similaritate structurală cu flavodoxinele mitocondriale(97) care conţin flavinmononucleotide şi au rol în transferul de electroni din procese metabolice microbiene(98). Un alt alergen fungic potenţial cross-reactiv de tip YCP4 este Cla h 7 din Cladosporium herbarum(12,21). Proteina YCP4 de tip flavodoxină are roluri în răspunsul la stres şi în reglarea genelor legate de metabolism(96).
Alt a 8 este manitol-dehidrogenaza (MtDH) dependentă de nicotinamid-adenin-dinucleotid-fosfat (NADP). Alt a 8 este un alergen minor de 29 kDa, cu secvenţă completă de 266 de aminoacizi(100). rAlt a 8 este recunoscut de anticorpii IgE din serurile a 41% dintre pacienţii cu alergie la Alternaria alternata(7,100). S-a dovedit reactivitatea încrucişată între Alt a 8 şi MtDH Cla h 8 din Cladosporium herbarum(12,100). MtDH este o enzimă implicată atât în sinteza manitolului, cât şi în degradarea prin catalizare a conversiei reversibile a manitolului în fructoză. Manitolul este cel mai comun poliol din fungi, constituind aproximativ 11% din greutatea uscată a Alternaria alternata, şi poate fi depistat în concentraţii mari atât în micelii, cât şi în spori. Deşi nu este necesar pentru germinarea sporilor, el serveşte drept carbohidrat de stocare sau de translocare(101,102). Localizarea alergenului Alt a 8 în compartimentele de tip vacuolar ale hifelor este în corelaţie cu funcţia sa biologică de MtDH. Alt a 8 este localizat numai în citoplasma hifelor, nu şi în spori(15).
O discuţie mai detaliată referitoare la alergiile alimentare la pacienţii sensibilizaţi la fungi merită avută în acest context. Anafilaxia la manitol, prezent în mod natural în ciupercile cultivate (Agaricus bisporus), dar şi în rodii (Punica granatum), poate fi asociată la acelaşi pacient cu anafilaxia ulterioară la manitol ca excipient medicamentos(103,104). Un astfel de caz a fost raportat la manitol din comprimate masticabile cu cisapridă(103). Au fost publicate de asemenea şi alte cazuri izolate de urticarie şi angioedem la manitol din granule efervescente cu paracetamol(105) şi de anafilaxie la manitol ca excipient din soluţia injectabilă de paracetamol(106,107). Deoarece manitolul este o haptenă inertă fără grupări reactive, s-a propus ca agent sensibilizant o bază Schiff a D-manozei formată prin reacţie Maillard(103). Referitor la asocierea posibilă între sensibilizarea la fungi aeropurtaţi precum Alternaria alternata şi unele alergii alimentare la ciuperci şi spanac(56), MtDH şi MnSOD din Agaricus bisporus au omologie semnificativă cu Alt a 8 şi, respectiv, Alt a 14, fiind de menţionat în cazul reacţiilor alimentare severe la ciuperci comestibile(12).
Alt a 9 este o proteină cu structură şi funcţie necunoscute. Greutatea moleculară a acestui alergen minor este de 43 kDa şi are relevanţă la doar 5% dintre pacienţii alergici(1). Alt a 9 este listat în baza de date Allergome, dar nu şi în baza de date a Subcomitetului OMS/IUIS pentru nomenclatura alergenelor(21).
Alt a 10 este o aldehid-dehidrogenază (ALDH) dependentă de nicotinamid-adenin-dinucleotid (NAD). Acest alergen minor de 53 kDa, cu secvenţă completă de 497 de aminoacizi, leagă IgE serice la doar 2% dintre pacienţii alergici la Alternaria(19). ALDH este implicată în catabolismul etanolului prin metabolizarea acetaldehidei(108). Un alt alergen potenţial cross-reactiv de tip ALDH este Cla h 10 din fungul Cladosporium herbarum(21).
Alt a 12 este o proteină P1 ribozomală acidică de 11 kDa. Similar cu Alt a 5, care este proteina P2 acidică 60S, Alt a 12 este o ribonucleoproteină fosforilată a cărei secvenţă completă cuprinde 110 aminoacizi. P1 şi P2 formează dimeri care au rol structural în subunităţile ribozomale mari şi participă direct la procesul de sinteză proteică(1). Alte alergene fungice de tip proteină P1 ribozomală sunt Cla h 12 din Cladosporium herbarum, Asp f 26 din Aspergillus fumigatus, Pen b 26 din Penicillium brevicompactum şi Pen ch 26 din Penicillium chrysogenum (sinonim: Penicillium notatum)(12,21).
Alt a 13 este o glutation-S-transferază (GST) de 26 kDa care reacţionează cu IgE serice de la 82% dintre pacienţii sensibilizaţi la Alternaria alternata(19), de aceea a fost considerat de unii specialişti alergen major, iar de alţii alergen minor, dar relevant(1,13). Alt a 13 are secvenţa completă de 231 de aminoacizi şi posedă capacitatea de a cataliza conjugarea formei reduse de glutation la substraturi xenobiotice în scop de detoxificare(109). Alergenul recombinant rAlt a 13 şi cel nativ nAlt a 13 au proprietăţi fizico-chimice, activitate enzimatică şi alergenicitate similare. rAlt a 13 are potenţialul de fi utilizat în diagnosticul molecular al cazurilor de alergie fungică(19,110). Alt a 13 este recunoscut ca alergen cu reactivitate încrucişată cu alte GST fungice precum Asp f GST din Aspergillus fumigatus, Cur l GST din Curvularia lunata, Cla h 13 din Cladosporium herbarum şi Epi p 13 din Epicoccum purpurascens(111).
Alt a 14 este enzima superoxid-dismutază dependentă de mangan (Alt MnSOD) localizată în principal la nivelul matricei mitocondriale(1,12). Ea a fost recunoscută recent ca alergen fungic minor, de 24 kDa(12,58). S-a constatat că această moleculă alergenică reacţionează cu anticorpii IgE din serurile pacienţilor sensibilizaţi la Alternaria alternata în proporţie de 11,5% şi i se pot atribui 6,6% dintre cazurile de alergie la acest fungus fără sensibilizare la alergenul major Alt a 1. Enzimele MnSOD conservate filogenetic sunt implicate în apărarea împotriva stresului oxidativ. O astfel de metaloenzimă converteşte radicalii superoxid în apă şi peroxid de hidrogen, care este apoi catalizat de către glutation peroxidază şi catalază în O2 şi H2O(1,12,38).
Este important de precizat că Alternaria alternata prezintă un nivel semnificativ de reactivitate încrucişată alergenică cu o serie de alte mucegaiuri, înrudite şi neînrudite filogenetic. Alt a 14 este, din această perspectivă, un panalergen din familia MnSOD. Din punctul de vedere al secvenţialităţii aminoacizilor, alergenele MnSOD din fungii Aspergillus fumigatus (Asp f 6), Malasezia sympodialis (Mal s 11), din latex Hevea brasiliensis (Hev b 10), fistic Pistacia vera (Pis v 4) sau de origine umană Homo sapiens (Hom s MnSOD) prezintă identităţi variind între 50% şi 88%(112). Alergenul Alt a 14 prezintă omologie semnificativă şi epitopi potenţial cross-reactivi cu Asp f 6. Dovezile de reactivitate încrucişată între Alt a 14 şi alergenul din Aspergillus fumigatus Asp f 6 în cazul aspergilozei bronhopulmonare alergice (ABPA) sugerează existenţa unui mecanism care poate fi legat de patogenia afecţiunii de hipersensibilitate(113), sensibilizarea mediată IgE faţă de biomarkerul ABPA Asp f 6 putând fi evaluată prin metode de imunoanaliză singleplex şi multiplex(114). S-a comentat că MnSOD ar putea avea implicaţii importante în sensibilizarea la Alternaria alternata ca factor de risc pentru dezvoltarea ABPA(12). Alt a 14 poate prezenta reactivitate încrucişată cu mangan-superoxid-dismutaza de origine umană Hom s MnSOD, care poate juca rol de autoalergen la un subgrup de pacienţi cu dermatită atopică sau eczemă nonatopică. Prin mimetismul molecular care duce la reactivitate încrucişată, o astfel de sensibilizare ar putea fi indusă primordial de expunerea la MnSOD fungică Mala s 11 a Malasezia sympodialis din microbiota cutanată(115). Se presupune că reacţiile mediate IgE la autoantigene sunt importante în perpetuarea simptomelor în afecţiunile alergice cronice, cum ar fi ABPA sau eczema atopică, şi în absenţa expunerii la alergene din mediu, evidenţiind relevanţa clinică a reactivităţii încrucişate(55). Mai mult, omologia dintre alergenele MnSOD din mucegaiuri (Alt a 14, Asp f 6) şi latex (Hev b 10), alături de cross-reactivitatea prin enolaze a condus la postularea posibilei existenţe a sindromului latex-mucegai(95,116).
Alt a 15 este un nou alergen minor de 58 kDa al Alternaria alternata, cu funcţie de serin-protează vacuolară. Anticorpii IgE serici de la 10,2% dintre pacienţii sensibilizaţi la Alternaria alternata leagă varianta recombinantă rAlt a 15(1). Alte alergene fungice de tip serin-proteazic cu potenţial cross-reactiv sunt: Cla h 9 din Cladosporium herbarum, Cla c 9 din Cladosporium cladosporioides, Asp f 13 şi Asp f 18 din Aspergillus fumigatus, Asp fl 13 şi Asp fl 18 din Aspergillus flavus, Pen ch 13 şi Pen ch 18 din Penicillium chrysogenum (sinonim: Penicillium notatum), Pen c2, Pen c 13 şi Pen ch 18 din Penicillium citrinum, Pen o 18 din Penicillium oxalicum, Rho m 2 din Rhodotorula mucilaginosa, Tri r 2 din Trichophyton rubrum şi Cur l 4 din Curvularia lunata(12,21). Toţi pacienţii cu IgE specifice faţă de rAlt a 15 sunt sensibilizaţi atât la Alternaria alternata, cât şi la Curvularia lunata, din cauza reactivităţii încrucişate extinse dintre serin-proteazele fungice. Diagnosticul molecular este util în cazurile cu sensibilizare la extract fungic natural de Alternaria alternata fără sensibilizare la alergenul major rAlt a 1, dar cu sensibilizare la alergenul cross-reactiv rAlt a 15. Această moleculă alergenică poate îmbunătăţi în viitor acurateţea diagnosticului şi managementul alergiei fungice mediate IgE(12).
Alt a 70 kDa este o glicoproteină cu greutate moleculară de 70 kDa care reprezintă 13% din greutatea uscată a extractelor naturale de Alternaria. Acest alergen cu funcţie biochimică necunoscută a fost raportat că induce rezultate pozitive la testarea cutanată la 87% dintre pacienţii sensibilizaţi la Alternaria alternata(117), în prezent fiind considerat alergen minor(13).
Alt a NTF2 este o proteină homodimerică de 28 kDa care reprezintă factorul 2 de transport nuclear, factor citozolic implicat în importul de proteine în nucleu prin complexul porului nuclear(1,118). NTF2 reprezintă un alergen minor cross-reactiv produs de Alternaria alternata şi Cladosporium herbarum, recunoscut şi de serurile de la pacienţii sensibilizaţi la Aspergillus fumigatus(119).
Alt a TCTP reprezintă o proteină tumorală controlată translaţional, cunoscută şi ca factor de eliberare a histaminei sau fortilină(120,121,122). TCTP este o proteină multifuncţională exprimată în toate organismele eucariote, care participă la progresia ciclului celular şi la proliferarea celulară, are proprietăţi antiapoptotice şi activitate de tip citokinic(122,123,124,125), cu acţiuni proinflamatoare şi de stimulare a eliberării de histamină în forma sa extracelulară dimerică(120,126,127). rAlt a TCTP a relevat o prevalenţă de aproximativ 4% a reactivităţii IgE (alergen minor) şi epitopi IgE cross-reactivi cu omologul Cla h TCTP al Cladosporium herbarum(128,129).
Diagnosticul molecular în alergia la Alternaria alternata
Aplicarea unui diagnostic alergologic molecular de precizie (PAMD@) este o abordare modernă, cunoscută anterior şi sub denumirea de diagnostic bazat pe componente alergenice(130). Componentele alergenice moleculare ale Alternaria alternata utilizate în PAMD@ în practica clinică sunt proteina fungică specifică rAlt a 1 şi enolaza fungică cross-reactivă rAlt a 6(57). Din datele ştiinţifice publicate în ultimul deceniu, se conturează propunerea ca şi alte molecule alergenice, inclusiv superoxid-dismutaza Alt a 14 şi serin-proteaza Alt a 15, să fie incluse în diagnosticul molecular al alergiei la Alternaria(1,38,131).
Metodele de analiză imunoenzimatică pentru IgE specifice faţă de componente alergenice din Alternaria alternata sunt de tip singleplex sau multiplex. Metoda singleplex de analiză imunoenzimatică cu componente alergenice cuplate la polimer celulozic activat ca fază solidă (capsulă) şi detectare fluorimetrică poate fi utilizată pentru determinarea IgE specifice serice faţă de cel mai important alergen molecular recombinant al fungusului Alternaria alternata, rAlt a 1(132). Metoda multiplex de analiză imunoenzimatică cu componente alergenice imobilizate în microarray pe lamă de sticlă preactivată ca fază solidă (chip) şi detectare fluorimetrică poate fi utilizată pentru determinarea în serul pacienţilor a IgE specifice faţă de două dintre alergenele moleculare recombinante ale Alternaria alternata: rAlt a 1 şi rAlt a 6(132). Metoda multiplex de explorare alergologică prin analiză imunoenzimatică cu componente alergenice cuplate cu nanoparticule activate depozitate în macroarray pe membrană de nitroceluloză ca fază solidă (chip) şi detectare colorimetrică poate fi utilizată pentru determinarea IgE specifice serice faţă de aceleaşi două componente alergenice moleculare recombinante importante ale Alternaria alternata: rAlt a 1 şi rAlt a 6(133).
Concluzii
Evaluarea detaliată a profilurilor moleculare de sensibilizare mediată IgE la fungusul Alternaria alternata folosind componentele sale alergenice recombinante este utilă în practica clinică pentru identificarea sensibilizării autentice faţă de cea cross-reactivă la pacienţii cu alergie la fungi. Cunoaşterea tuturor alergenelor moleculare ale Alternaria alternata este utilă clinicianului pentru a înţelege factorii trigger şi mecanismele imunologice complexe implicate în alergia la mucegaiuri şi pentru a-l familiariza cu noi biomarkeri diagnostici potenţiali care să îmbunătăţească în viitor acurateţea diagnosticului şi managementul alergiei fungice mediate IgE.
Alternaria alternataalergene moleculare