Comparing oxidative metabolism in patients with uveal melanoma versus cutaneous melanoma
Comparaţie între metabolismul oxidativ la pacienţii cu melanom uveal versus melanom cutanat
Abstract
Introduction. Cutaneous and uveal malignant melanomas (MM) show different metastasis characteristics, with no approved systemic treatment for metastatic uveal melanoma. Melanocytes are particularly susceptible to oxidative stress, defined as an imbalance between the formation of prooxidants and their removal by antioxidant systems. A better understanding of oxidative stress markers (lipid peroxides, albumin thiols, total antioxidants) differences between cutaneous and uveal melanoma could be useful in quantifying the different response to treatment. Materials and method. In a prospective study, 45 patients diagnosed with cutaneous MM (PC), as well as a cohort of 44 patients diagnosed with uveal MM (PU) were enrolled and separated into three distinct subgroups according to the received treatment: radiotherapy (14), enucleation (18), enucleation with postoperative systemic chemotherapy. All patients were followed-up for two years after diagnosis, obtaining seven determinations of biochemical parameters of oxidative stress – markers of lipid peroxidation inferement via malondialdehyde measurement, total thiol-albumin groups (Shoshinsky method), for the inferement of oxidative proteic degradation. Results. All levels were elevated compared to baseline in cutaneous and uveal melanoma patients: serum lipid peroxides PC 8.229, PU 8.377 versus baseline 4 µmol/100 ml, serum albumin thiols 511.8 PC, PU 479.9 versus baseline 450 µmol/L, serum total antioxidants 2.768 PC, 3.040 PU versus baseline 1.2 µmol/L, suggesting oxidative stress increase in the presence of a malign tumor. The levels in uveal melanoma patients varied more compared to those from cutaneous melanoma. Conclusions. The results indicate an increase of lipid peroxidation reaction, serum albumin thiols and serum total antioxidants, in correlation with tumor evolution, both in uveal and cutaneous melanoma compared to baseline values. As such, the presence of the tumor is an oxidative stress inductor, with anti-tumor treatment also having the ability to induce oxidative stress through the destruction of tumor cells. Changes between the two groups indicate different molecular signaling mechanisms of endogenous antioxidant activity. They are continuously increasing in cutaneous melanoma, however in uveal melanoma, at the end of the treatment, there is a decrease of recorded serum levels of oxidative parameters. It is thus suggested that uveal melanoma leads to the installation of a more pronounced oxidative stress compared to cutaneous melanoma, with an intensification of oxygen metabolism and a lower response capacity of the body, possibly related to the increased aggressiveness of this malign cell type. This could explain the increased resistance to immunotherapy of uveal melanoma when compared to cutaneous melanoma. Our results can be useful in monitoring the tumor evolution under treatment.Keywords
malignant uveal melanomalipid peroxidationtotal antioxidantsRezumat
Introducere. Melanoamele maligne (MM) cutanate şi uveale prezintă caracteristici diferite de metastazare, fără un tratament sistemic standardizat pentru melanomul uveal metastatic. Melanocitele sunt deosebit de susceptibile la stresul oxidativ, definit ca un dezechilibru între formarea prooxidanţilor şi îndepărtarea acestora de către sistemele antioxidante. O mai bună înţelegere a diferenţelor markerilor de stres oxidativ (peroxizi lipidici, tioli de albumină, antioxidanţi totali) dintre melanomul cutanat şi cel uveal ar putea fi utilă în cuantificarea răspunsului diferit la tratament. Materiale şi metodă. Într-un studiu prospectiv, 45 de pacienţi diagnosticaţi cu melanom malign cutanat (PC), precum şi o cohortă de 44 de pacienţi diagnosticaţi cu melanom malign uveal (PU) au fost înrolaţi şi împărţiţi în trei subgrupe distincte, în conformitate cu tratamentul primit: radioterapie (14), enucleare (18), enucleare cu chimioterapie sistemică postoperatorie. Toţi pacienţii au fost urmăriţi timp de doi ani după diagnostic, obţinându-se şapte determinări ale parametrilor biochimici ai stresului oxidativ – markeri de deducere a peroxidării lipidelor prin măsurarea malondialdehidei, grupe tiol-albumină totală (metoda Shoshinsky), pentru deducerea degradării proteice oxidative. Rezultate. Toate nivelurile au fost crescute faţă de valoarea iniţială la pacienţii cu MM cutanat şi uveal: peroxizi lipidici serici PC 8,229, PU 8,377 faţă de valoarea iniţială de 4 µmol/100 ml, tioli de albumină serică 511,8 PC, PU 479,9 faţă de valoarea iniţială de 450 µmol/l, antioxidanţi serici totali 2,768 PC, 3,040 PU faţă de valoarea iniţială de 1,2 µmol/L, sugerând o creştere a stresului oxidativ în prezenţa unei tumori maligne. Concluzii. Nivelurile parametrilor de stres oxidativ la pacienţii cu MM uveal au variat mai mult decât în cazul MM cutanat şi au evoluat atât în MM uveal, cât şi în cel cutanat, în comparaţie cu valorile iniţiale. Ca atare, prezenţa tumorii este un inductor de stres oxidativ, tratamentul antitumoral având, de asemenea, capacitatea de a induce stres oxidativ prin distrugerea celulelor tumorale. Modificările din cele două grupuri indică diferite mecanisme de semnalizare moleculară a activităţii antioxidante endogene. Ele cresc continuu în MM cutanat, însă mai accentuat în MM uveal. La sfârşitul tratamentului, se observă o scădere a nivelurilor serice înregistrate ale parametrilor oxidativi. Se sugerează astfel că MM uveal duce la instalarea unui stres oxidativ mai pronunţat comparativ cu melanomul cutanat, cu o intensificare a metabolismului oxigenului şi o capacitate de răspuns mai scăzută a organismului, posibil legată de agresivitatea crescută a respectivului tip de celule maligne. Acest fapt ar putea explica rezistenţa crescută la imunoterapie a MM uveal în comparaţie cu MM cutanat. Rezultatele noastre pot fi utile în monitorizarea evoluţiei tumorii sub tratament.Cuvinte Cheie
melanom malign uvealperoxidare lipidicăantioxidanţi totaliIntroduction
Melanocytes are located in various parts of the human body, such as the skin and the eye(1). Their transformation leads to melanoma, an aggressive and deadly neoplasm(1). Cutaneous and uveal melanomas show different characteristics, including significant differences in genetic alterations, metastatic sites and therapeutic response(1). In recent decades, great efforts have been made to obtain a more comprehensive understanding of genetics, genomics and molecular changes, enabling the identification of key cellular processes and signaling pathways in melanomas(1). Major breakthroughs were realized in the treatment of metastatic cutaneous melanoma, but most patients relapse(1). Currently, there is no approved systemic treatment for metastatic uveal melanoma(1). Thus, these two different cancers are in therapeutic need to overcome treatment failure and improve patients’ prognosis(1).
Malignant melanoma, a neoplasm arising from malignant transformation of melanocytes, is predominantly a disease of the skin, but in rare instances it may occur at other sites, including the mucous membranes (hard palate, maxillary gingiva, lip, throat, esophagus, vulva, vagina and perianal region) and the eye (uvea and retina)(2).
Melanocytes are particularly susceptible to oxidative stress owing to the prooxidant state generated during melanin synthesis and to the intrinsic antioxidant defenses that may be shattered in pathologic conditions(2).
Oxidative stress can be defined as an imbalance between the formation of prooxidants and the ability of the body antioxidant systems to decrease or remove their harmful effects(3). This redox state may result in disruption of redox signaling and biomolecules damage(3).
Oxidative stress can disrupt the homeostasis of melanocytes, causing damage to DNA, protein and cellular components(2). Altered reactive oxygen species (ROS) levels could also affect epigenetic mechanisms and promote alterations in gene expression, thus leading to severe impairment of cell survival and to cancer development(2). Reactive oxygen species are highly reactive molecules that are constantly produced in all aerobic organisms, mostly as a consequence of aerobic respiration(4). The term covers several types of chemical species of short-lived molecules with unpaired electrons, including free radicals such as superoxide (O2-) or hydroxyl (-OH), and nonradicals such as hydrogen peroxide (H2O2)(4). Levels of reactive oxygen species (ROS) are reduced by antioxidant defenses, but increased by transition metals such as iron or copper and by exogenous agents such as ionizing radiation or ozone(4).
Similarly, nitrogen-derived free radicals are called reactive nitrogen species (RNS) and their utmost representative precursors are nitric oxide (NO) and peroxynitrite (ONOO-)(4). Nitric oxide (NO) is well known to be a product of the catalytic action of the nitric oxide synthase (NOS) enzyme family on L-arginine(4).
The main endogenous sources of ROS in mammalian cell include: mitochondria, endoplasmic reticulum, peroxisomes, cytosol, plasma membrane, and extracellular space(3). The exogenous sources of ROS are related to the physical factors exposure, such as UV solar light, gamma rays radiation exposition, electromagnetic field, alpha particles emitted by radioactive elements decay, environmental chemicals exposure (excessive pollution by organochlorines, xenobiotics, aromatic amines, quinones, polycyclic aromatic hydrocarbons, ozone, singlet oxygen [1O2], pathogens, chemotherapy, tobacco smoking, iron overload, trauma, drugs and physical exercise)(3).
A cell functioning is dependent on its redox state, that is the ratio of the reversible oxidized molecules and the reduced form of a specific redox couples in a cell, such as oxidized/reduced glutathione (GSSG/GSH), nicotinamide adenine dinucleotide (NAD) cation/reduced NAD (NAD+/NADH) or nicotinamide adenine dinucleotide phosphate (NADP) cation/reduced NADP (NADP+/NADPH) and the balance between oxidants formation and protection by antioxidants systems(3).
The link between cancer metabolism and redox homeostasis
It has been shown that melanoma cells regularly reprogram their metabolism to provide an equivalent reduction and support in antioxidant protection(5). There are different signaling pathways involved in supplying and regulation redox power in melanoma cells(5). In particular, it has been demonstrated that during melanomagenesis the oxidative pentose phosphate pathway (ox-PPP), serine biosynthesis and 1-CM (one-carbon metabolism) are responsible for adenine dinucleotide phosphate (NADPH) and glutathione (GSH) production(5). On the other hand, monocarboxylate transporters (MCTs) and glycolytic enzymes, such as pyruvate kinase (PK)-M2 (PKM2), are also associated to redox homeostasis during melanoma initiation and progression(5).
Pentose phosphate pathway (PPP) is primarily catabolic and serves as an alternative glucose oxidizing pathway for the generation of NADPH that can be involved in redox metabolic adaptation of melanoma(5). The antioxidant role of glucose-6-phosphate dehydrogenase (G6PD), the rate-limiting enzyme of PPP that catalyzed the first reaction with the production of NADPH, has been studied in an in vitro melanoma model(5). Indeed, the inhibition of G6PD sensitized malignant melanoma cells A375 to oxidative stress, decreased proliferation and induced apoptosis(6).
Another study showed that the high expression of G6PD promotes melanoma growth via the signal transducer and activator of transcription 3/5 (STAT3/5) pathway in a human melanoma xenograft model(7). The role of G6PD in cooperation with NADPH oxidase 4 (NOX4) for the support of redox homeostasis has been related to melanoma cells in vitro, indeed targeting both enzymes suppressed cell proliferation(8). These studies indicate that PPP represents an essential redox metabolic pathway in melanoma and serves as a pivotal role in survival and adaptation of melanoma cells.
Glycolysis is a metabolic pathway converting glucose into pyruvate and lactate as final metabolites, and releasing energy to form ATP and NADH molecules. Several enzymes belonging to this metabolic pathway have been found to be associated with melanoma genesis(5).
A unique biochemical feature of the melanocyte is the generation of melanin, which leads to the generation of hydrogen peroxide and the consumption of reduced glutathione (GSH)(10). Successful attempts to reverse or inhibit this process have been initiated(9), but to date they have not been clinically tested.
Melanoma cells overexpress a redox-depending enzyme, PKM2, an isoform of the pyruvate kinase, that converts phosphoenolpyruvate into pyruvate, being the last irreversible reaction of aerobic glycolysis(11). Reactive oxygen species oxidize a specific cysteine residue in PKM2, thus diverting glucose away from lactate production and towards the oxidative branch of pentose phosphate pathway, leading to increased NADPH production and, thus, to redox homeostasis(5). These data provide a direct link between cancer metabolism and redox homeostasis. Melanoma cell invasion and metastasis levels were positively correlated with high PKM2 activity as well as the glycolytic capability. Knockdown of PKM2 markedly attenuated the malignant phenotypes of melanoma cells, including cell proliferation, invasion and metastasis in vitro and in vivo, suggesting that PKM2 is a potential therapeutic target in melanoma(11).
Metastatic pathways in patients with melanoma
Metastasis represents the end-product of an intricate biological process, which necessarily involves dissemination of neoplastic cells to different anatomic sites and adaptation of neoplastic cells to foreign tissue microenvironments(12).
The process of metastasis is determined by the interplay between metastatic tumor cells, various host factors and homeostatic mechanisms(13). Metastasis is a multistep process, which includes proliferation, neovascularization, immune system evasion, lymphangiogenesis, invasion, circulation, embolism, extravasation and colonization(13).
The interactions between the neoplastic cells and the non-neoplastic stromal cells are important in the progression of the invasion-metastasis cascade(12,14).
Cutaneous melanoma can metastasize hematogenously or lymphogenously(15). The three predominant models that endeavor to explain the patterns of melanoma progression are the stepwise spread model, the simultaneous spread model and the model of differential spread(15). The stepwise spread model posits that melanoma metastasizes initially via the lymphatic system towards regional lymph nodes and, subsequently, systemic dissemination occurs(16,17).
The second predominant model is the simultaneous spread model, which involves that primary cutaneous melanoma metastasizes simultaneously by hematogenous and lymphatic pathways(18).
The third model, which attempts to explain the patterns of progression of cutaneous melanoma, has been coined the model of differential spread(17). This model proposes that there are multiple independent dissemination pathways(17).
The time course to the development of metastases differs between the different metastatic routes(15). There are several clinical and histopathological risk factors for the different metastatic pathways(15). In particular, patient’s sex and the anatomical location of the primary tumor influence the patterns of disease progression(15).
Metastatic disease is detectable at diagnosis in less than 4% of uveal melanoma cases(19). Uveal melanoma metastasizes exclusively through hematogenous dissemination of tumor cells preferentially and almost exclusively to the liver, with up to 90% of metastatic uveal melanoma associated with hepatic lesions, and in most cases the liver is the only affected organ(20,21). Other sites, such as the lungs (24%), bones (16%), skin/subcutaneous tissues (11%) and lymph nodes (10%), can also be affected(21), while the involvement of brain and fellow eye is rare.
In this paper, we undertook a comparative analysis of the evolution of markers of oxidative stress (lipid peroxides, albumin thiols, total antioxidants) in patients with cutaneous malignant melanoma compared to those with uveal melanoma. The aim is to describe possible differences in oxygen metabolism between the two diseases, differences that may be clinically useful or may justify the relatively different biological behaviors of these two locations or, potentially, the different response to systemic treatments.
Materials and method
In this study, 45 patients, respectively 19 women and 26 men, aged between 30 and 84 years old, with cutaneous malignant melanoma, as well as a cohort of 44 patients diagnosed with uveal malignant melanoma were enrolled. Due to the heterogeneity of the patients’ group, in terms of age, sex and the extent of the disease at initial presentation, the patients with uveal melanoma were separated into three distinct subgroups, depending on the therapeutic method applied, respectively:
-
the first group was treated with radiotherapy (n=14);
-
the second group was treated strictly by enucleation (n=18);
-
the third group was also treated by enucleation, but with systemic treatment applied postoperatively (mainly chemotherapy, only one case with interferon immunotherapy) (n=12).
All patients were followed-up clinically and biologically for two years after the diagnosis, respectively before and after the initial medical intervention, and subsequently at regular intervals of 3-6 months. Seven determinations of biochemical parameters of oxidative stress were obtained:
-
markers of lipid peroxidation by measuring the reaction of malondialdehyde (MDA) as a final product;
-
total thiol-albumin groups (by the Shoshinsky method), as a measure of the oxidative degradation of circulating proteins.
Results
For all patients, a series of biochemical tests were performed that followed oxygen metabolism and the installation of oxidative stress in the dynamics of the treatment. The therapeutic approach was chosen in accordance with established protocols that targeted optimal tumor regression, patient’s quality of life and overall health. Lipid oxidation parameters were initially analyzed in order to identify changes that could lead to demonstrable differences in the mechanisms of development of the two types of melanoma, cutaneous versus uveal. The average values obtained by measuring the final concentration of MDA, a product of lipid degradation under the action of reactive oxygen species, are presented in Figure 1. For reference, the baseline value was 4 µmol/100 ml.
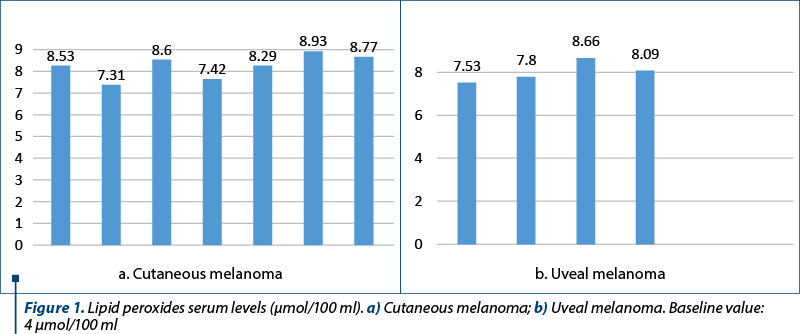
The recorded values show fluctuations due to the treatment applied, with overall increase in the level of lipid peroxidation. Oxidative stress can affect all the organic molecules of the cell, which become targets for the following damage: deoxyribonucleic acid (DNA), proteins, lipids, carbohydrates. For effectively following the therapeutic intervention, the initial target of oxidative stress must be known. For example, DNA is known to be the initial target of damage produced by the addition of H2O2 in mammalian cell cultures, such that DNA strand breaks occur before “detectable” lipid peroxides or “detectable” oxidized proteins. The method of detecting lesions produced on some target molecules can give incomplete information; thus, highlighting protein damage through the detection of carbonyl radicals in the initial stages of the damage can be negative, but the determination of the oxidation of -SH groups, which occurs earlier, is positive. Trying to apply the results from the experimental model to the patients enrolled in the study, we investigated the biochemical parameters of oxidative stress in the dynamics of the applied treatment, considering that they can provide additional data related to the metabolism of the malignantly transformed cell, data that correlate with the angiogenesis process and, last but not least, data related to the destructive evolution of malignant cells, following equally the effectiveness of the treatment. Following the dynamics, it is observed in all investigated patients the increase in the intensity of the reaction measured, suggesting, in accordance with literature data, that the presence of a tumor is associated with an increase in oxidative stress, and this is why surgical excision can decrease oxidative stress. It is important to take into consideration that lipid peroxidation is a reaction with a chain mechanism; the measured effects are total, even those at a distance, thus suggesting an increase in the concentration of the final product measured.
For providing a more comprehensive picture, albumin thiols were also measured, reflecting the intensity of protein degradation under the attack of reactive oxygen radical species (baseline value: 450 µmol/L). The results (Figure 2) show the increase of the oxidative attack on the proteins as well, suggesting the installation of oxidative stress.
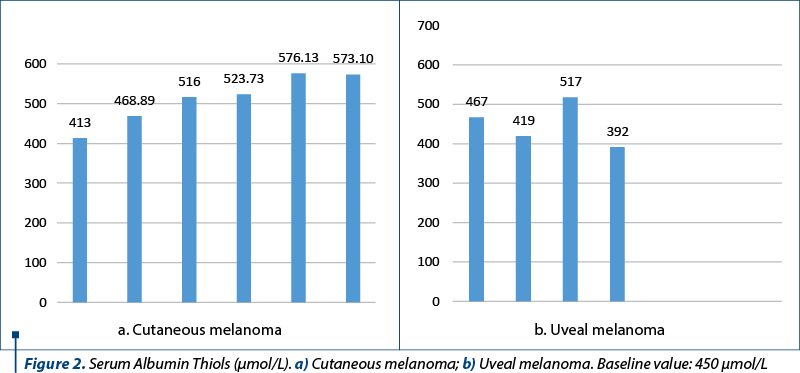
Comparing the oxidative degradation of proteins by measuring the end-product, namely albumin thiols, changes are observed between the two cohorts. The values are higher in the case of patients with uveal melanoma, once again confirming the hypothesis that in uveal melanoma there is a higher oxidative stress than the one which occurs after the treatment of cutaneous melanoma.
Next, the values of total antioxidants were measured, which, through the values recorded and presented in Figure 3, suggest the activation of natural antioxidant protection systems in response to an installed oxidative stress (baseline value: 1.2 µmol/L). It is difficult to say whether this increase is due only to endogenous antioxidants, because no study or questionnaire on the intake of exogenous antioxidants has been done.
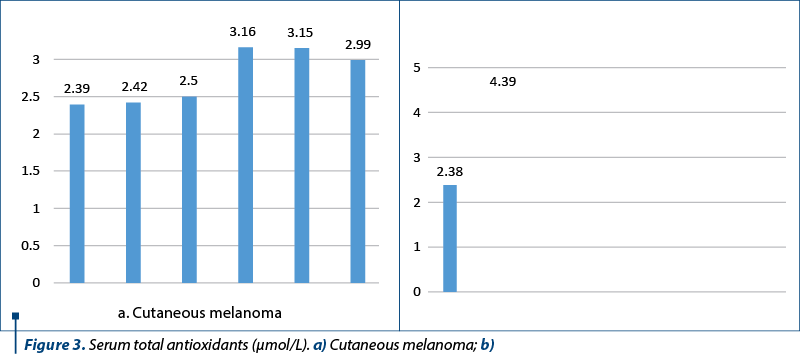
We consider that each patient is his own witness, and the recorded values are related to the initial value for each patient. Furthermore, the study assumed that there were no essential lifestyle changes during the investigations (no changes were reported by the patients). We could draw the conclusion that the tumor is an oxidative stress factor to which the body reacts at the cellular level. The investigated parameters show values consistent with the state of health and could be used in patient monitoring and treatment management.
In cancer, the tumor presence itself is an oxidative stress inductor; however, anti-tumor treatments also have the ability to induce oxidative stress, by destroying tumor cells through degradative processes initiated at the molecular level.
Changes between the two groups indicate different molecular signaling mechanisms of endogenous antioxidant activity. They are continuously increasing in cutaneous melanoma; however, in uveal melanoma, at the end of the treatment, there is a decrease of recorded serum levels of oxidative parameters. It is thus suggested that the uveal melanoma leads to the installation of a more pronounced oxidative stress compared to cutaneous melanoma, with an intensification of oxygen metabolism and a lower response capacity of the body, possibly related to the increased aggressiveness of this malignant cell type. This could explain the increased resistance to immunotherapy of uveal melanoma when compared to cutaneous melanoma.
Discussion
Reactive oxygen species behave as a double-edged sword at the cellular level. When reactive species of oxygen are in a low concentration, they have a role in cell signaling, converting the cell phenotype to angiogenesis and neovascularization, which contributes to tumor growth and proliferation. On the other hand, when ROS production is in excess, it becomes destructive and damages tumor cells. The generation of oxidative stress during the process of tumor growth and development is particularly highlighted by the stimulation of lipid peroxidation, a process that generates numerous electrophilic aldehydes, stable compounds that can diffuse inside the cell, exerting several cytotoxic effects. In addition, lipid peroxidation products could be responsible for blocking cell cycle and/or apoptotic mechanisms, leading to decreased therapeutic efficacy and the establishment of treatment resistance. Excess lipid peroxidation products can inhibit cell cycle progression by blocking cyclin-dependent kinases, keeping tumor cells in the G0 phase, prolonging the G1 phase, and delaying entry into the S phase, reducing the rate of tumor cell proliferation.
Although intense oxidative stress could be effective in inducing tumor cytotoxicity, moderately induced oxidative stress could have opposite effects, contributing to the development of resistance mechanisms, decreasing therapeutic efficacy. Moreover, numerous in vivo and in vitro studies(22,23) claim that tumor cells and tissues suffer an intense, chronic, sublethal oxidative stress, compared to normal cells, even in the absence of prior exogenous stimulation, being absolutely necessary for tumor viability and growth. Intense intratumorally oxidative stress may play an important role in the process of tumor invasion and metastasis.
The action of active oxygen metabolites at the level of structural or enzymatic proteins causes their denaturation(3,10). A critical factor in this oxidation reaction of protein SH groups by ROS is the steric factor. Exposure of thiol groups in proteins is essential. This explains the increased sensitivity to oxidation of denatured proteins (this property is used as an effective method for identifying the content of thiol groups, as well as for identifying the secondary and tertiary structure of proteins). By oxidative degradation, enzymatic activity is inhibited, as a result of the modification of the active catalytic center (due to the oxidation of thiol groups or hemeprotein, but also by oxidative degradation of the aromatic cycles in the structure of some amino acids). Structural proteins are inactivated due to changes induced by polymerization reactions, branching or induced cleavage of polypeptide chains by ROS.
In many human conditions, oxidative stress is only a consequence, not a cause of the pathological disease process. Tissue damage by aggressive agents, such as infections, trauma, toxins, extreme temperatures etc., leads to the overproduction of “injury mediators”, such as prostaglandins, leukotrienes, interleukins and cytokines (such as tumor necrosis factor – TNF), which play an important role in the production of tissue lesions. In the neoplastic cell, the establishment of an imbalance between the production of reactive oxygen species and the elimination of free radicals leads to a state of oxidative stress and to the destruction of some essential components of the cell. Several studies(4,10,23) concluded that oxidative stress resulting from this imbalance has a causative role but is also a consequence of carcinogenesis, interfering with all phases of this process, such as initiation, promotion, progression, invasion and metastasis.
In the patients enrolled in the study, we investigated the biochemical parameters of oxidative stress in the dynamics of the applied treatment, considering that they can provide additional data related to the metabolism of the malignantly transformed cell, data to correlate with the angiogenesis process and, finally, data related to the destructive evolution of malignant cells, equally aiming at the efficiency of the treatment. Lipid peroxidation reaction is a reaction with a chain mechanism, the measured effects are total, even those at a distance, thus suggesting an increase in the concentration of the final product measured. Since this parameter alone cannot provide a complete picture, we also measured albumin thiols, a measure of the intensity of protein degradation under the attack of reactive oxygen radical species. Following the dynamics, an increase in the intensity of the measured reaction is observed in all investigated patients, suggesting, as the data are also presented in the specialized literature(24,25), that the presence of a tumor is associated with an increase in oxidative stress, while surgical resection could induce its decrease. Finally, it is worth noting that proteins are not the primary target of oxidative attack.
Conclusions
The results indicate an increase of lipid peroxidation reaction, serum albumin thiols and serum total antioxidants in correlation with tumor evolution, both in uveal and cutaneous melanoma, in comparison with baseline values. As such, the presence of the tumor is an oxidative stress inductor, with anti-tumor treatment also having the ability to induce oxidative stress through the destruction of tumor cells. Changes between the two groups indicate different molecular signaling mechanisms of endogenous antioxidant activity. They are continuously increasing in cutaneous melanoma; however, in uveal melanoma, at the end of the treatment, there is a decrease of recorded serum levels of oxidative parameters. It is thus suggested that uveal melanoma leads to the installation of a more pronounced oxidative stress compared to cutaneous melanoma, with an intensification of oxygen metabolism and a lower response capacity of the body, possibly related to increased aggressiveness of this malignant cell type. This could explain the increased resistance to immunotherapy of uveal melanoma when compared to cutaneous melanoma. Our results can be useful in monitoring the tumor evolution under the treatment.
Conflicts of interests: The authors declare no conflict of interests.
Bibliografie
-
Bertolotto C. Cutaneous and uveal melanoma: two different cancers in therapeutic needs. C R Biol. 2021;344(3):219-231.
-
Venza M, Visalli M, Beninati C, De Gaetano GV, Teti D, Venza I. Cellular Mechanisms of Oxidative Stress and Action in Melanoma. Oxid Med Cell Longev. 2015;2015:481782.
-
Kruk J, Aboul-Enein HY, Kładna A, Bowser JE. Oxidative stress in biological systems and its relation with pathophysiological functions: the effect of physical activity on cellular redox homeostasis. Free Radic Res. 2019;53(5):497-521.
-
Ortega AL, Mena S, Estrela JM. Oxidative and nitrosative stress in the metastatic microenvironment. Cancers (Basel). 2010;2(2):274-304.
-
Arslanbaeva LR, Santoro MM. Adaptive redox homeostasis in cutaneous melanoma. Redox Biol. 2020;37:101753.
-
Li D, Zhu Y, Tang Q, et al. A new G6PD knockdown tumor-cell line with reduced proliferation and increased susceptibility to oxidative stress. Cancer Biother Radiopharm. 2009;24(1):81-90.
-
Hu T, Zhang C, Tang Q, et al. Variant G6PD levels promote tumor cell proliferation or apoptosis via the STAT3/5 pathway in the human melanoma xenograft mouse model. BMC Cancer. 2013;13:251.
-
Cai T, Kuang Y, Zhang C, et al. Glucose-6-phosphate dehydrogenase and NADPH oxidase 4 control STAT3 activity in melanoma cells through a pathway involving reactive oxygen species, c-SRC and SHP2. Am J Cancer Res. 2015;5(5):1610-1620.
-
Jenkins NC, Grossman D. Role of melanin in melanocyte dysregulation of reactive oxygen species. Biomed Res Int. 2013;2013:908797.
-
Obrador E, Liu-Smith F, Dellinger RW, Salvador R, Meyskens FL, Estrela JM. Oxidative stress and antioxidants in the pathophysiology of malignant melanoma. Biol Chem. 2019;400(5):589-612.
-
Zhou Y, Huang Z, Su J, et al. Benserazide is a novel inhibitor targeting PKM2 for melanoma treatment. Int J Cancer. 2020;147(1):139-151.
-
Gupta GP, Massagué J. Cancer metastasis: building a framework. Cell. 2006;127(4):679-695.
-
Fidler IJ. The biology of melanoma metastasis. J Dermatol Surg Oncol. 1988;14(8):875-881.
-
Valastyan S, Weinberg RA. Tumor metastasis: molecular insights and evolving paradigms. Cell. 2011;147(2):275-292.
-
Adler NR, Haydon A, McLean CA, Kelly JW, Mar VJ. Metastatic pathways in patients with cutaneous melanoma. Pigment Cell Melanoma Res. 2017;30(1):13-27
-
Mervic L. Time course and pattern of metastasis of cutaneous melanoma differ between men and women. PLoS One. 2012;7(3):e32955.
-
Tejera-Vaquerizo A, Barrera-Vigo MV, Fernández-Canedo I, et al. Estudio temporal de los diferentes patrones metastásicos en la progresión del melanoma cutáneo [Longitudinal study of different metastatic patterns in the progression of cutaneous melanoma]. Actas Dermosifiliogr. 2007;98(8):531-538.
-
Pizarro Á. Models of melanoma spread and final results of the Multicenter Selective Lymphadenectomy Trial-I. Actas Dermosifiliogr. 2015;106(2):82-85.
-
Finger PT, Kurli M, Reddy S, Tena LB, Pavlick AC. Whole body PET/CT for initial staging of choroidal melanoma. Br J Ophthalmol. 2005;89(10):1270-1274.
-
Collaborative Ocular Melanoma Study Group.. Assessment of metastatic disease status at death in 435 patients with large choroidal melanoma in the Collaborative Ocular Melanoma Study (COMS): COMS report no. 15. Arch Ophthalmol. 2001;119(5):670-676.
-
Diener-West M, Reynolds SM, Agugliaro DJ, et al. Development of metastatic disease after enrollment in the COMS trials for treatment of choroidal melanoma: Collaborative Ocular Melanoma Study Group Report No. 26. Arch Ophthalmol. 2005;123(12):1639-1643.
-
Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020;38(2):167-197
-
Aboelella NS, Brandle C, Kim T, Ding ZC, Zhou G. Oxidative Stress in the Tumor Microenvironment and Its Relevance to Cancer Immunotherapy. Cancers (Basel). 2021;13(5):986.
-
Bisevac JP, Djukic M, Stanojevic I, et al. Association Between Oxidative Stress and Melanoma Progression. J Med Biochem. 2018;37(1):12-20.
-
Budirincă MA, Gruia MI, Murgoi G, Anghel R. Monitoring oxidative status in patients with uveal melanoma during oncological treatment. Oncolog-Hematolog.ro. 2019 Dec;49(4):32-5.

A teaching experience in geriatric oncology: the Treviso SIOG advanced courses
Silvio Monfardini
Cancer predominantly affects older patients, but an integrated approach bringing together clinical oncologists and geriatricians for the best management of neoplasia in the elderly is not worldwide available. There is a need for some kind of shortcut that would teach the principles of geriatric oncology w...
Variability of response to multimodal therapy in a stage IV bronchial carcinoid: a case report
Alexandru Ioan Băjenaru, Laura Predescu, Mirela Gherghe
Neoplasmele pulmonare neuroendocrine (NEN) se clasifică în tumori neuroendocrine (NET) şi carcinoame neuroendocrine (NEC). NET cuprind tumorile carcinoide tipice (TC) şi atipice (AC), în timp ce NEC sunt clasific...
Real-world and clinical trials for assessing the effectiveness of new drugs in oncology. The role of observational studies with exemplification in non-small cell lung cancer (NSCLC)
Mihaela Teodorescu, Alexandru Grigorescu
Datele din lumea reală (real-world data; RWD) şi studiile observaţionale (OS) sunt din ce în ce mai importante în cercetarea ...
Sialic acid as a tumoral marker in uveal melanoma
Mihai Adrian Păsărică, Christiana Diana Maria Dragosloveanu, Paul Filip Curcă, Cosmin-Ionuţ Nisipaşu, Alina-Gabriela Gheorghe, Alexandru-Călin Grigorescu
Melanomul uveal este cea mai frecventă tumoră intraoculară, cu o rată mare de mortalitate din cauza metastazelor, în special a celor hepatice....
Ultrasonography in necrotic choroidal melanoma
Mihai Adrian Păsărică, Christiana Diana Maria Dragosloveanu, Paul Filip Curcă, Alina Ciocâlteu, Alexandru C. Grigorescu
Melanomul uveal este cea mai frecventă tumoră malignă intraoculară la adulţi, cu originea în tractul uveal al globului ocular. Necroza spontană a melanoamelor coroidale este foarte rară, dar unele melanoame uveale po...