Consensul celor 6 sexe. De la biochimie la comportament
The consensus of the 6 sexes. From biochemistry to behaviour
Abstract
The sex of a newborn is stated at birth. Then, the presence of external sexual organs expresses the sex: male or female. It was stated that the native sex is the “principal sex”, with his “sexual character”.The evident sex follows a process which takes place in consensus: from the genetic sex (XX or XY) to pubertary sex, or adult sex.
Therefore, the sex is made through 6 steps: genetic sex, gonadal sex, the sex of genital internal organs, the sex of external genital organs, the neuro-comportamental (or hypothlamico-cerebral sex) and the pubertary sex or “scondary” sex.
This sex classification in 6 analytical stages starts from the observation of sex daviations. The are diseases of the genetic sex (genetic anomalies, eg. Turner syndrome), diseases of the gonads (hermaphroditism), anomalies of internal and/or sexual organs (pseudohermaphroditism). Recently, the troubles of sexual behaviour could be considered as pseudohermaphroditism, because there is missing the consensus generated by what is considered as conventional normality in biology (the average plus/minus 2 standard deviations). The lack of pubertary sexual development (late puberty) or precocious development complete the consensus of the 6 sexes.
Keywords
main sexual charactersecondary sexual charactergonadeneurobehavioural sexsexual orientationgen identityRezumat
Sexul unui copil nou născut se afirmă la naştere. Atunci, prezenţa caracteristică a organelor „genitale externe“ exprimă sexul: feminin sau masculin, femeiesc sau bărbătesc. S-a afirmat astfel că sexul nativ este „sexul principal“ cu „caracterul său sexual principal“.Sexul aparent este urmarea unui proces ce se desfăşoară în consens: de la sexul genetic (XX sau XY) la cel pubertar (sau al adultului).
Astfel, sexul se formează prin 6 trepte: sexul genetic, gonadic, al organelor genitale externe, al organelor genitale interne, sexul neuro-comportamental (sau hipotalamo-cerebral) şi sexul pubertar sau „secundar“.
Împărţirea/clasificarea sexului în 6 trepte analitice pleacă de la observaţiile asupra devierilor de la sex. Există boli ale sexului genetic (anomalii genetice, ex. sindrom Turner), boli ale gonadelor (hermafroditismul adevărat), anomalii ale organelor genitale, interne şi/sau externe (pseudohermafroditism). Mai recent, anomaliile comportamentului sexual pot fi considerate forme de pseudohermafroditism, pentru că lipseşte consensul generat convenţinal de „normalul“ în biologie (media plus/minus 2 deviaţii standard). Lipsa de dezvoltare sexuală la pubertate (întârziată) sau dezvoltarea precoce întregesc „consensul“ celor 6 sexe.
Cuvinte Cheie
caracter sexual principalcaracter sexual secundargonadesex neurocomportamentalorientare sexualăidentitate de genI. Introducere. Normalul în biologie
Convenţional, se consideră normalul în biologie, pentru datele continue (cele care sunt măsurabile, de exemplu glicemia, colesterolemia, înălţimea, greutatea, testosteronemia, estradiolemia etc.) ca media plus/minus două deviaţii standard (SD). Pe baza acestor date se construieşte o curbă Gauss-Laplace, de tipul „clopot“.
Pentru datele discontinui, care sunt entităţi şi nu măsuri, este dificil a trasa limite convenţionale. De aceea, multe din datele discontinui sunt interpretate pe baza unor teste de tip X2 (hi pătrat) sau z (zed), cu ajutorul unor grupe de control. Limita este dată tot de o convenţie, aceea de a genera o semnificaţie statistică de p < 0,05. De exemplu, la un lot de analiză de 1000 de persoane, pentru a spune că un fenomen este patologic, el trebuie observat la 4 persoane (0,4%), cu condiţia ca în lotul de control fenomenul să nu apară (se generează un X2 de 4,01, cu p = 0,0453).
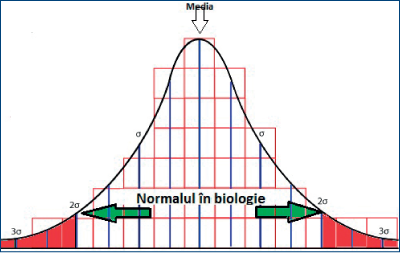

Comportamentul sexual reprezintă o dată discretă. El nu poate fi definit prin măsurători, ci prin comparare.
II. Consensul celor 6 sexe
Sexul unui copil nou-născut se afirmă la naştere. Atunci, prezenţa caracteristică a organelor „genitale externe“ exprimă sexul: feminin sau masculin, femeiesc sau bărbătesc. S-a afirmat astfel că sexul nativ este „sexul principal“, cu „caracterul său sexual principal“.
Statistic vorbind, marea majoritate a celor care posedă sex feminin la naştere vor deveni femei în viaţa adultă, iar marea majoritate a celor care posedă sex masculin la naştere vor deveni bărbaţi în viaţa adultă. Ia naştere, astfel, „caracterul sexual secundar“.
Dar nu toţi cei cu un caracter „sexual principal“ evoluează spre un caracter „sexual secundar“ conform cu evoluţia considerată normală. Medicina cunoaşte destule devieri patologice de la acest traseu. Cea mai cunoscută cauză a fost castrarea băieţilor pentru a deveni „eunuci“. Chiar dacă cercetarea ştiinţifică nu exista în acel moment, era evident că testiculul generează caracterul sexual „secundar“ masculin. Testiculul a fost prima „gonadă“ descrisă ca atare. Ulterior, la sexul feminin, gonada a fost identificată prin ovar.
O altă veche „cunoştinţă“ de nerealizare uzuală a sexului secundar este produsul Hermafroditos, fiul de sex bărbătesc al lui Hermes şi al Afroditei, cu nimfa de sex femeiesc Salmacis. Produsul format astfel avea amândouă sexele. El a generat chiar termenul medical de „hermafroditism“, de la care a plecat, apoi, cel de „pseudohermafroditism“.
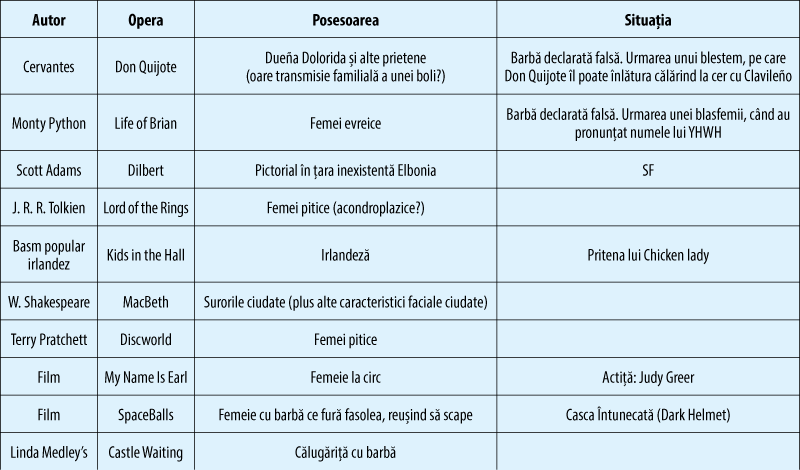
Mult mai recent, observaţiile clinice au arătat că, la „pubertate“, o parte din băieţi devin femei şi unele fete devin bărbaţi, sau cu compontente importante ale celuilalt sex. De exemplu, la pubertate, la unii băieţi apare ginecomastia (creşterea sau apariţia unui sân feminin), iar la unele fete apare şi se dezvoltă barba (există vestitele femei cu barbă – tabelul 1, figura 1).
William Shakespeare a scris nu numai despre femei cu barbă, dar şi despre homosexuali. Un celebru pasaj din „Neguţătorul din Veneţia“ sugerează (la 1550) că dragostea nu trebuie neapărat să treacă prin inimă, ci se poate şi prin cap**. Astăzi, fenomenul este clar: la nivel cerebral se integrează comportamentele sexuale.
Plecând de la aceste constatări timpurii (de peste 3 secole de medicină observaţională), s-a căutat cauza.

În acel moment, s-au pus în evidenţă relaţiile dintre sexul vizibil şi organele interne legate de sex. Ele au fost numite organe „genitale interne“. Tot atunci, s-a conturat noţiunea de „caracter sexual terţiar“, legat de aspectul sexual al vârstnicilor: bărbaţii vârstnici capătă caractere feminine, iar femeile vârstnice capătă caractere masculine.
Medicina a evoluat prin observaţii clinice şi experimente. Observaţiile clinice au scos în evidenţă o pleiadă de anomalii ale organelor genitale externe şi interne, numite hermafroditism sau pseudohermafroditism. Experimentele au încercat să găsească cauza. Astfel, hermafroditismul a fost definit prin prezenţa a două gonade diferite (testosteron şi ovar) sau prin prezenţa de celule gonadice diferite (ovar cu celule din testicul – celule Leydig – sau testicul cu foliculi ovarieni). Pseudohermafroditismul a fost definit ca modificări de intersexualitate, dar cu o gonadă clar constituită şi unică. Boala apare fie prin hormonii celuilalt sex în exces, fie prin lipsa de receptivitate a testosteronului sau a hormonilor testiculari (vezi mai departe) la nivelul organelor genitale interne şi/sau externe.
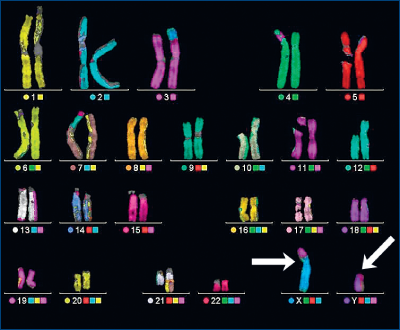
La sfârşitul veacului al XIX-lea, s-au izolat celulele cu nucleii lor. Ulterior, în nuclei s-au izolat cromozomii. Omul are 46 de cromozomi, din care 2 sunt puternic implicaţi în dezvoltarea sexului. Astfel, s-au descris 44 de cromozomi somatozomi şi 2 cromozomi sexuali.
Cu alte cuvinte, sexul este dependent de 2 cromozomi, denumiţi „cromozomi sexuali“, numiţi X şi Y. Organismele feminine au 2 X, cele masculine au 1 X şi 1 Y. Pentru un om sănătos se scrie: 44 XX pentru femeie şi 44 XY pentru bărbat.
Analiza cromozomilor, la pacienţi cu diferite tulburări în sfera organelor sexuale (genitale interne şi externe), precum şi a caracterelor sexuale secundare, a adus surprize nemaiîntâlnite în planul evoluţiei fenotipice a organismelor. De atunci, s-a observat că modificări ale cromozomilor (cel mai frecvent X şi Y, dar şi alţii) conduc la modificări în sfera sexuală; se modifică expresia normală a gonadelor, a organelor genitale interne şi/sau externe, a caracterului sexual secundar (pubertar).
În centrul verigilor patogenice, de la alcătuirea cromozomilor şi până la apariţia manifestărilor de boală, au fost izolaţi „hormonii“. Ei au primit numele de „hormoni sexuali“ şi au fost clasificaţi pe trepte de comandă, astfel: hormoni hipotalamici, hipofizari şi gonadali. Clasificarea este de fapt inversă cu descoperirea lor. Primii hormoni sexuali, izolaţi la sfârşitul secolului al XIX-lea, au fost cei gonadali, apoi hormonii gonadotropi hipofizari au fost observaţi în anii ’20-’30 ai secolului XX, iar hormonii hipotalamici, releasing şi inhibiting pentru hipofiză, au fost puşi în evidenţă în anii ’60.
Ultima achiziţie importantă legată de sex a avut loc în anii ’80 ai secolului XX. S-a observat că centrii hipotalamici releasing, responsabili de secreţia hipofizară de gonadotropi (stimulatori ai gonadelor), au caracter funcţional ciclic la femei şi aciclic la bărbaţi. Acest tip de comportament hormonal a fost denumit sex neurocomportamental. Termenul comportamental a fost adăugat pentru că, pe lângă secreţiile neuronale identificate, s-a observat că structura sexuală respectivă (hipotalamusul) integrează şi unele comportamente specifice sexului secundar (voi reveni).
Cele mai recente cercetări asupra relaţiilor neurocomportamentale folosesc acum tehnici avansate de rezonanţă magnetică nucleară şi de tomografie cu emisie de pozitroni. Prin aceste tehnici, zone din creier pot fi „vizualizate“ în cursul stimulărilor sexuale, fie cu feromoni sau prin filme video.
Astfel, s-a conturat ideea de organizare în compartimente a sexului. S-au formulat cele 5 compartimente ale sexului, care determină 6 niveluri de sexualizare. Se ştia că există relaţii temporale între unele compartimente descrise: vezi dependenţa caracterului sexual terţiar de cel secundar, la rândul lui dependent de cel primar. Ştiinţa a reuşit să identifice şi celelalte relaţii temporale dintre toate componentele sexului.
Ierarhia componentelor sexului
-
Sexul cromozomial
-
Sexul gonadal
-
Sexul organelor genitale interne
-
Sexul organelor genitale externe
-
Sexul neurocomportamental
-
Sexul secundar (pubertar)
Atunci a apărut ideea de consens al componentelor sexului. Consensul este realizat atunci când sexul generator primordial este acelaşi cu caracterul sexual secundar şi terţiar.

Important pentru psihiatrie este „orientarea“ sexuală, generată de sexul neurocomportamental.
Nivelul unu al sexualizării:
sexul cromozomial
Factorul generator al sexului organismului este clar definit prin „sexul genetic“. La sfârşitul veacului al XIX-lea, s-au izolat celulele cu nucleii lor. Ulterior, în nuclei s-au izolat cromozomii. Omul are 46 de cromozomi, din care 2 sunt puternic implicaţi în dezvoltarea sexului. Astfel, s-au descris 44 de cromozomi somatozomi şi 2 cromozomi sexuali. Celula ce conţine XX este femeie, celula ce conţine XY este bărbat.
Acest sex se numeşte „sex primordial“. El este sexul cromozomial.
S-a observat că sexul primordial nu formează sexul secundar sau terţiar. El nu determină nici sexul organelor genitale interne. Singura acţiune sexuală a sexului primordial, a cromozomilor sexuali, este de a dezvolta gonada, adică „sexul gonadal“ sau „primitiv“. Cuplul XY dezvoltă testicul, iar cuplul XX dezvoltă un ovar normal.
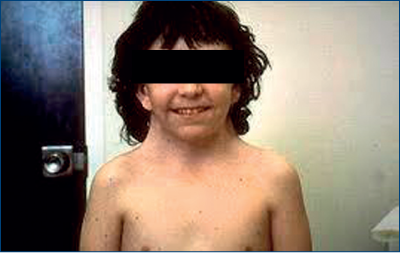
Lipsa cromozomilor sexuali conduce la apariţia unor tulburări în sfera sexuală. Cazul clasic este cel al sindromului Turner.
Tulburarea esenţială este legată de lipsa activităţii ovarului (plus statura mică).
În acest sindrom, genotipul (suma cromozomilor) este 45 X0. Adică există lipsa unui gonozom, fie el Y sau X.
Interesant este că aceşti pacienţi, care ar trebui să fie „unisex“, sunt, fenotipic (ca aspect exterior), femei***. În plus, aceşti pacienţi, de fapt paciente, prezintă uter, ovare, trompe. Dar ovarele sunt nefuncţionale.

Acest fenomen arată că tiparul (pattern-ul) după care sunt alcătuite organismele vertebrate sexuate pe Pământ este de tip feminin . Pe tiparul femeii, „cineva“ a intervenit şi a creat bărbatul sau sexul bărbătesc. Ştiinţa este, în acest punct, împotriva afirmaţiei că „mai întâi a fost bărbatul (Adam)“ şi că femeia a apărut din „coasta acestuia“. În plus, din acest fapt, i.e., că fenotipul unui organism X0 este feminin, rezultă şi că manifestarea sexului este subînţeleasă a fi de tip feminin, iar că, pentru a te manifesta masculin, trebuie să se întâmple nişte evenimente dependente de sexul masculin, adică subsecvente existenţei cromozomului Y. Sau, cu alte cuvinte, dacă în evoluţia intrauterină (aşa cum arată studiile mai vechi) şi/sau imediat prenatal (aşa cum arată studiile mai noi) nu există acţiunea testosteronului, comportamentul sexual va fi de tip feminin.
Există situaţii mai rare în care un cromozom Y alterat nu reuşeşte să genereze un testicul în integralitatea sa. Atunci, în organismul respectiv, ce apare mai ales ca femeie (acelaşi tipar…!), se observă o gonadă mixtă, un ovotestis, conglomerat testicular într-un ovar. Sindromul se numeşte hermafroditism adevărat.
Dovada este certă: pentru a fi bărbat, trebuie ca mai întâi să fii femeie!
Nivelul doi al sexualizării: sexul gonadal
După constituirea sexului cromozomial, care produce o gonadă normală, se observă un fenomen diferit la sexul bărbătesc faţă de cel femeiesc. În timp ce ovarul, cu cei 150.000 de foliculi, rămâne în repaus până la 10-11 ani, testiculul devine activ în momentul formării.
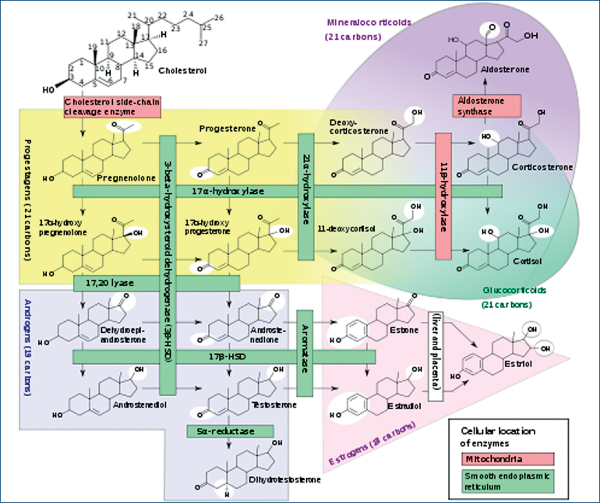
Din momentul producerii intrafetale a gonadei masculine, aceasta preia acţiunea de realizare a consensului sexual. Intrauterin, în lunile 2-3, testiculul secretă cantităţi uriaşe de testosteron, de 100 de ori mai mari decât la bărbatul adult (convenţional, testosteronemia adultului este de 10-30 nmol/l). El are rolul de a determina formarea organelor genitale interne („sexul primar“) şi a organelor genitale externe („sexul secundar“) specifice sexului masculin.
Organele sexuale interne sunt reprezentate de prostată şi veziculele seminale. Organul sexual extern este penisul. Acestea toate sunt generate prin acţiunea de transformare locală a testosteronului în dihidrotestosteron.
Testosteronul gonadal, prin DHT, stimulează formarea organelor genitale interne („sexul primar“). Organele sexuale interne sunt reprezentate de prostată şi veziculele seminale.
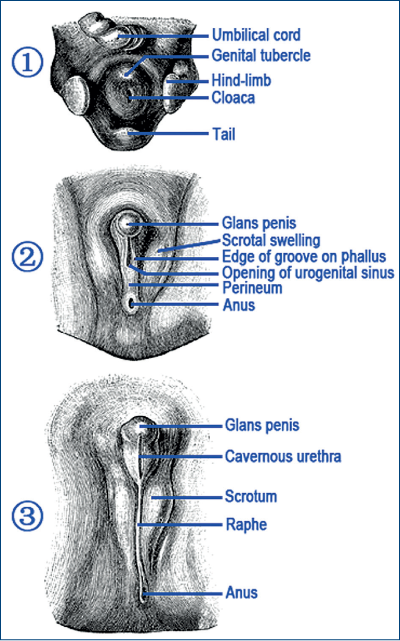
Testosteronul gonadal, tot prin DHT, stimulează mugurele sexual, care va genera organul sexual extern, penisul, adică „sexul secundar“.
Testosteronul gonadal, tot prin DHT, stimulează mugurele sexual, care va genera organul sexual extern, penisul, adică „sexul secundar“.
Acţiuni ale testosteronului
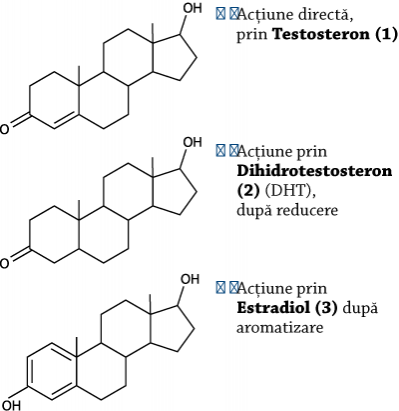
Sediile principale de acţiune şi urmări
1) Testicul ➙ Prima acţiune: stimulează paracrin spermatogoniile şi spermiogeneza
1) Muşchi ➙ Stimulează intrarea glucozei în miocite prin mecanisme insulin-independente; urmarea: hipoglicemie ➙ scade hiperinsulinemia ➙ reduce fenomenul de downregulation
1) Ficat ➙ Stimulează formarea de IGF-I, în prezenţa GH ➙ creşte eficacitatea insulinei
2) Ţesuturi sexual-dependente ➙ Prostata, firul de păr, pielea
3) Creier ➙ Stimulează libidoul; în creier, 70% din testosteron se transformă în estradiol
Testiculul mai secretă doi hormoni. Primul este Mullerian inhibitory factor – MIF sau anti mullerian hormone (AMH), în cantităţi, de asemeni, uriaşe. El are rolul de a distruge formarea organelor genitale interne specifice sexului feminin (generate din aparatul primordial Mullerian).
Al doilea este „megalina“, care are rolul de a „sparge“ ţesuturile şi a permite testiculului să coboare din regiunea renală, unde ia naştere, în afara corpului, în scrot. Motivul este că testiculul adultului produce testosteron numai sub 33°C, şi nu la 37°C, cât există în interiorul organismului.
Se observă, de fapt, că, intrauterin, numai gonada masculină funcţionează, cea feminină nu are nici o acţiune. Consensul XX din celula feminină face ca organismul feminin să fie mult mai uşor de realizat intrauterin, faţă de lupta „intrinsecă“ X-Y în celula masculină. Încă o dovadă a tiparului feminin al organismului uman (şi nu numai).
Ovarul începe să funcţioneze la pubertate. El va secreta estrogeni şi progesteron, cu rol de pregătire a uterului pentru momentul nidării unui ou/zigot. Interesant este că estrodiolul, hormonul feminin ce produce proliferare endometrială uterină, se formează în ovar prin trecerea de la colesterol la testosteron şi, apoi, prin aromatizare, la estradiol. Din acest punct de vedere se poate afirma că „pentru a fi femeie trebuie să fii bărbat“!


Nivelurile trei şi patru ale sexualizării: organele genitale
Lipsa unei cantităţi adecvate de testosteron la nivelul mugurelui organului genital extern, lipsa de AMH sau de megalină determină diferite forme de boli sexuale ale sexului bărbătesc. Aceste boli, cu testicul, au primit numele de pseudohermafroditism masculin (tabelul 3) pentru a le diferenţia de hermafroditismul adevărat, unde gonada este mixtă. În aceste cazuri, pseudohermafroditismul masculin înseamnă lipsa parţială sau totală a caracterelor sexuale masculine şi/sau prezenţa de elemente sexuale feminine la un organism genetic sexual masculin (XY) şi cu testicul (Pentru a complica lucrurile, există şi pseudohermafroditism invers: fete cu ovare au organe genitale masculine; de obicei, boala apare prin leziuni de glande suprarenale).
Pseudohermafroditismul masculin (sursa: Pereţianu, 1985)
I.
Ireceptivitate testiculară la LH/hCG; Sindrom de testicule afuncţionale (sd. Berthesène)
II.
Defecte enzimatice de sinteză
a testosteronului
a) Deficit dublu: testicul şi corticosuprarenală
-
Deficit 20-22 desmolază – OGE sunt feminine
-
Deficit 3 beta ol-hidroxi-steroid-dehidrogenaza (sd. Bongiovani)
-
Deficit de 17‑hidroxilază (sd. Biglieri)
b) Deficit unic testicular
-
Deficit de 17-20 desmolază (sd. Zachman)
-
Deficit de hidroxi-steroid-oxidoreductaza (sd. Saez-Reifenstein)
III.
Deficite ale receptorului de androgeni
a) Testiculul feminizat complet (sd. Morris – se prezintă ca cele mai frumoase femei şi se manifestă ca atare) sau cu variantele incomplete.
b) Deficit de 5-alfa-reductază: lipsa transformării T în DHT (lipsa organelor genitale externe, declarate fete la naştere, hiperandrogenism pubertar prin testicul ectopic).
IV.
Deficit de MIF: generează persistenţa canalelor mulleriene (feminine)- (sd. Royer)
V.
Disgenezii testiculare de cauze multiple; sd. de disgenezie testiculară, sd. Gordon-Overstreet
VI.
Ingestie maternă de estrogeni (alimentaţie, toxice, disruptori – vezi mai departe)
Aceste anomalii cuprind o arie largă de forme clinice, de la lipsa penisului şi a scrotului (A) la lipsa numai a scrotului (B) sau la incompleta dezvoltare a organelor genitale externe (C) (figura 13).
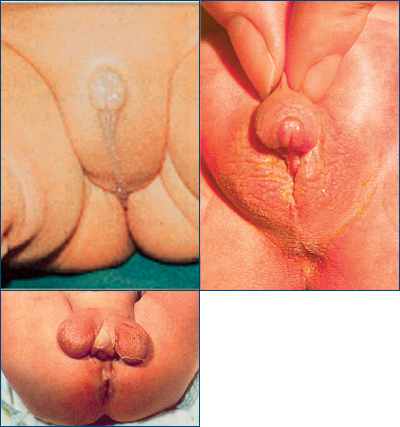
Cele mai frecvente modificări de pseudohermafroditism sunt hipospadias (un orificiu anormal localizat la nivel ureteral pe faţa ventrală a penisului – boală de care a suferit Ludovic al XVI-lea al Franţei) şi criptorhidismul (lipsa testiculului/lilor din scrot şi prezenţa lui/lor pe traiectul descendent, de obicei în canalul inghinal).
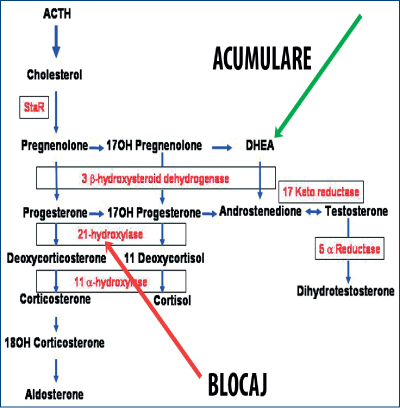
La fete, există pseudohermafroditism prin androgenizarea organelor sexuale externe. Fenomenul apare cel mai frecvent în deficitul de 21-hidroxilază corticosuprarenală în hiperplazia congenitală de suprarenală. Deficitul formării de cortizol în fasciculată conduce la creşterea ACTH, care produce hiperplazia reticulatei, cu hiperproducţie de androgeni (DHEA). De vină este tot „steroidogeneza“.
Nivelul al cincelea al sexualizării: sexul neurocomportamental
Sexul neurocomportamental este încă un element important în structuralizarea sexului. Producerea lui are loc tot intrauterin, între săptămânile 8 şi 24, cel mai frecvent între lunile a 4-a şi a 5-a (Hines, 2006). Testiculul, prin testosteron, pune pecetea asupra sexului neurocomportamental: prezenţa testosteronului în hipotalamusul fătului în lunile 4 şi 5 va conduce la comportamentul aciclic al hipotalamusului sexual, prin aciclicitatea GnRH.
Intrauterin, în lunile 4 şi 5, se poate spune că toate celulele organismului fetal bărbătesc înoată în testosteron. Testosteronemia fătului este de peste 100 de ori mai mare ca a celui mai activ bărbat adult!
Dacă nu există îndeajuns de mult testosteron în hipotalamus sau dacă testosteronul ajunge acolo în cantităţi adecvate de‑abia în luna a 6-a intrauterin (adică tardiv), sau dacă nu există îndeajuns de mulţi receptori pentru acţiunea specifică a testosteronului, atunci hipotalamusul devine ciclic, cu evidenţierea ciclică a hipofizei sexuale, prin gonadotropi. Este un element important pentru diferenţierea bolilor legate de comportamentele adulte ale bărbaţilor transsexuali şi homosexuali.
Aceste date sunt certe: creierul unui băiat va fi de la naştere diferit de cel al unei fetiţe. El este impregnat testosteronic. Se ştie că un astfel de creier se va comporta la nivel hipotalamic sexual pe linie masculină. Ceea ce încă nu se ştie este dacă şi alte zone cerebrale se vor comporta diferit, adică bărbătesc vs. femeiesc, indiferent de contextul social.
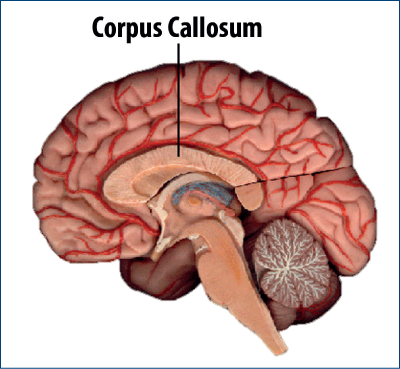
Cercetări recente sugerează că există dimorfism sexual în dezvoltarea a numeroase componente legate de comportamentul sexual: hipotalamus, cortex, nucleul median amigdalian, cunoscute componente cu numeroşi receptori pentru testosteron. De asemenea, un rol important pare a‑l avea corpul calos, cel care face legătura dintre cele două emisfere (Manhães, 2002).
Cercetările moderne demonstrează că există un dimorfism funcţional diferit al corpului calos la femei vs. bărbaţi: creierul stâng este cel primordial la femei, iar transferul informaţiilor se face stânga-dreapta, în timp ce la bărbaţi este invers (Yakota, 2005).
Nivelul mic de testosteron intrauterin conduce, la băieţi, la comportamente postnatale specifice fetiţelor (Hines, 2006).
Ipotetic, dar nedovedit clinic, este că se poate ca anumite cantităţi de testosteron ovarian să poată ajunge în lunile 4-5 în hipotalamusul embrionului feminin. Dacă este aşa, dar faptele nu sunt dovedite, este posibil ca, la maturitate, femeia să aibă comportamente de tip masculin. Cert este că acest comportament nu blochează ciclicitatea hipotalamo-hipofizară generatoare a ciclului ovarian de care depinde ciclul uterin/menstrual.
Argumente pentru susţinerea tezei comportamentului masculin prin androgenizare intrauterină sunt susţinute şi de observaţiile asupra bolnavelor cu hiperplazie congenitală de corticosuprarenală, o boală cunoscută cu creştere intrauterină de androgeni. Aceste fete se comportă ca băieţii.
Cercetările recente arată că nu numai testosteronul intrauterin este important, dar şi hipertestosteronemia tranzitorie postnatală, la 3-4 luni, este importantă pentru definirea ulterioară a comportamentului de tip masculin (Alexander, 2014). În plus, s-a sugerat şi că anumiţi factori hormonali peripubertari pot avea importanţă (Berenbaum, 2011).
În acest sens, cercetări pe şoricei arată că manipularea estrogenică/antiestrogenică peripartum la şoricei este importantă. Ea conduce la comportamente homosexuale la sexul bărbătesc, dacă se administrează antiestrogeni peripartum (He, 2013). În schimb, administrarea de estrogeni creează, paradoxal, comportamente puternic masculine!! Mecanismele nu depind de aromatizarea testosteronului, ci de acţiunea directă a estrogenilor.
Nivelul al şaselea de sexualizare
„Ultimul sex“ este cel secundar. Este vorba despre dezvoltarea sexuală la pubertate. Ea depinde de secreţia testosteronului, la băiat, şi de cea estrogenică şi progestativă, la fete.
Leziuni ale acestui sex sunt destul de frecvente. Cel mai des este observată întârzierea apariţiei pubertăţii. Mai demult, cea mai cunoscută cauză a fost castrarea băieţilor pentru a deveni „eunuci“.

Chiar dacă cercetarea ştiinţifică nu a existat în acel moment, era evident că testiculul generează caracterul sexual „secundar“ masculin.
Pubertatea tardivă este definită ca lipsa caracterelor sexuale secundare la 18 ani. Sindromul a primit numele de hipogonadism. Cauzele sunt legate de leziuni gonadice (hipogonadism primar), hipofizare (hipogonadism secundar), hipotalamice (hipogonadism terţiar) sau receptoriale (hipogonadism cuaternar). Cel mai frecvent, boala are cauză hipotalamică, prin lipsă de maturizare a axei, care poate fi deblocată, paradoxal, cu hormoni ai gonadelor care nu funcţionează, testosteron pentru băieţi şi estradiol pentru fete. Dar paradoxul nu există, pentru că exact aceşti hormoni, şi mai ales testosteronul, generează niveluri adecvate de aromatază cerebrală (Cornill, 2011).
O altă veche „cunoştinţă“ de nerealizare uzuală a sexului secundar este produsul Hermafroditos, fiul de sex bărbătesc al lui Hermes şi al Afroditei, cu nimfa de sex femeiesc Salmacis. Produsul format astfel avea amândouă sexele.

El a generat chiar termenul medical de „hermafroditism“, de la care a plecat, apoi, cel de „pseudohermafroditism“.
Actualitatea în acest domeniu o reprezintă identificarea a ceea ce se numeşte „endocrine disruptors“. Este vorba despre micropoluanţi, despre care s-a arătat că modifică statistic important apariţia pubertăţii, nu numai la băieţi, dar şi la fete (Den Hond, 2006).
Privit din toate aceste puncte de vedere, pentru un clinician este mult mai uşor de identificat natura unei boli de sex, de clasificat hermafroditismul şi tulburările de comportament sexual. Tot astfel, este mai uşor de tratat hipogonadismul secundar sau terţiar, ca patologie a sexualizării caracterului sexual secundar şi cel cuaternar legat de social (caracterul sexual terţiar este cel de vârstă, când bărbaţii se feminizează, iar femeile se masculinizează).
La aceste caractere sexuale, unii adaugă şi sexul cuaternar, un epifenomen socio-cultural şi educaţional, pe care unii nu îl consideră ca element de sexualizare. Mecanismele legate de reproducrerea speciei sunt totuşi hormonale (Keefe, 2002).
III. Comportamentul sexual
Urmarea sexualizării corecte este comportamentul sexual. El este definit fiind consensual pentru cele 4 componente ale sale: dorinţă, orientare, activitate şi orgasm.

A. Dorinţa
Dorinţa sexuală se defineşte ca libidou. Acesta depinde, aproape în întregime, de nivelul de testosteron, atât la bărbat, cât şi la femeie. Lipsa dorinţei, a libidoului, conduce la neorientare sexuală.
Libidoul este generat la nivel cerebral, dar mecanismul depinde de capacitatea celulei nervoase de a prelua testosteronul pe cale aromatizării, spre estradiol, şi nu pe cea descrisă la nivelul organelor sexuale interne şi externe, a dihidrogenării, spre dihidrotestosteron (Morris, 2004). Reducerea aromatizării la nivel cerebral este implicată şi în tulburări cognitive la bărbat (Weinstock, 2011).
De aceea, se poate afirma, şi la acest aspect, că pentru a fi bărbat, trebuie să fii femeie. Adică pentru a te comporta ca un bărbat, cu libidou, orientare şi activitate adecvată, trebuie ca testosteronul, hormon masculin, să se transforme în celula cerebrală în estradiol, hormonul feminin. Interesant este însă că şi libidoul feminin este dependent tot de transformarea testosteronului în estradiol. De aceea, şi femeia are nevoie de testosteron.
Cu toate că femeia are niveluri serice de 10 ori mai mici de testosteron faţă de bărbat (1-2 nmol/l), acestea sunt mai mari decât nivelul de estradiol (0,5-1,2 nmol/l).
Testosteronul reprezintă cheia libidoului şi a comportamentului sexual normal al bărbatului adult. De nivelul de testosteron depind (Mogoş, 2004):
-
dorinţa sexuală şi excitaţia sexuală;
-
poluţiile nocturne;
-
tumescenţa peniană nocturnă, erecţii nocurne şi matinale spontane;
-
frecvenţa masturbaţiei;
-
frecvenţa coitală;
-
frecvenţa gândurilor şi fanteziilor legate de sex;
-
creşterea numărului de ejaculări, a lichidelor formatoare a spermei (ex: lichid seminal);
-
orientarea (driving) comportamentului spre sexul opus (heterosexualitate);
-
stimularea comportamentului sexual cu şi fără parteneră;
-
creşterea calităţii orgasmului, prin amplificarea contractilităţii musculaturii netede a glandelor accesorii şi distensia bulbului uretral.
O serie de parametri sociali pot modifica acţiunea şi/sau nivelul de testosteron. Printre aceştia au fost citaţi (Mogoş, 2004):
-
interacţiunea cu partenera creşte secreţia/concentraţia de testosteron;
-
bărbaţii care sunt implicaţi într-o relaţie stabilă au testosteronul mai redus decât cei care nu au parteneră (dar care sunt în căutarea uneia);
-
nivelul testosteronului este mai crescut în timpul weekend-ului;
-
testosteronemia creşte la bărbaţii care îşi doresc un copil, în perioada fertilă a partenerei;
-
anticiparea unei activităţi sexuale poate creşte nivelul testosteronului.
Activitatea sexuală reduce tranzitoriu numărul de receptori androgeni la nivelul creierului, printr-un fenomen de downregulation, fapt care se poate corela cu inhibiţia activităţii sexuale ce caracterizează saţietatea sexuală.
Se admite că nu există corelaţii între nivelul testosteronului şi excitabilitatea obţinută prin expunerea la material erotic explicit, cu toate că s-a arătat că poate reduce perioada de latenţă pentru apariţia excitaţiei la material erotic (Mogoş, 2004).
B. Orientarea sexuală
Orientarea sexuală (driving) sau preferinţa pentru un anumit tip de partener sexual este urmarea unui libidou normal, i.e. existent; lipsa libidoului conduce la neorientare sexuală!
Istoric, se afirmă că este normal ca bărbaţii să fie orientaţi sexual spre femei, iar femeile spre bărbaţi. Fenomenul defineşte heterosexualismul. Orientarea sexuală diferită de „consensul celor 6 sexe“ defineşte homosexualitatea. Tradiţional, cercetările bio-medicale asupra homosexualităţii s-au aplecat asupra atributelor biologice feminine pentru a caracteriza homosexualii bărbaţi şi, în antiteză, de cercetare a atributelor masculine, pentru a caracteriza femeile homosexuale (Gooren, 2011).
Această abordare nu este pe de-a-ntregul corectă. Ea poate fi combătută de observaţia clinică. Bărbaţii homosexuali se comportă homosexual în ceea ce priveşte orientarea (proiectată asupra altor bărbaţi). Acţiunea sexuală concretă este în fond bisexuală, dar numai cu alţi bărbaţi homosexuali.
Privit din acest punct de vedere, cum pot doi bărbaţi homosexuali să-şi producă (provide) satisfacţia sexuală dacă amândoi sunt orientaţi feminin, atât din punctul de vedere al spiritualităţii relaţiei, cât şi al repertoriului feminin al actului sexual, în acelaşi timp, din moment ce unul din cei doi se manifestă cu o orientare masculină şi cu un comportament masculin tipic?
Această perspectivă ridică întrebări asupra definirii orientării homosexuale în context hormonal, i.e. testosteronic. Comportamentul sexual feminin pare a nu fi dependent de testosteron, ca atare, el pare a fi dependent de construcţia specifică a unor arii cerebrale plecând de la pattern-ul feminin de bază. Dacă femeile prezintă comportamente homosexuale, atunci este posibil să fi fost influenţate în timpul dezvoltării intrauterine de nivelul lor endogen de testosteron (Ponseti, 2007). Faptul derivă din observaţia că femeile homosexuale prezintă mai puţină substanţă cenuşie la nivelul cortexului peririnal faţă de femeile heterosexuale (vezi mai departe).
C/D. Activitatea şi orgasmul vor fi abordate cu altă ocazie. Ele se realizează, la bărbat, prin erecţie şi ejaculare. La femeie, activitatea se realizează preferenţial pasiv, în relaţia cu activitatea bărbatului. Orgasmul este fie clitoridian (65%), fie vaginal (35%).
IV. Tulburări ale orientării de sex
A. Masculin
Homosexualitatea masculină este prezentă de când lumea, din moment ce este puternic criticată în vechi scrieri ale omului (vezi Vechiul Testament din Biblie). Care este cauza? Este ea de ordin social, sau este de ordin hormonal (Balthazart, 2011)?
Cercetări ştiinţifice asupra determinismului biologic al comportamanetului sexual de tip homosexual au debutat în anii ’70 ai secolului XX, prin observaţia princeps legată de „berbecul homosexual“. Berbecul homosexual este unul din puţinele animale disponibile care prezintă în mod natural (adică neinduse de experimente umane) comportamente homosexuale human-like. 8% dintre berbeci sunt homosexuali (Roselli, 2009, vezi şi figura 18). Nu se cunoaşte comportament lesbian la oi.
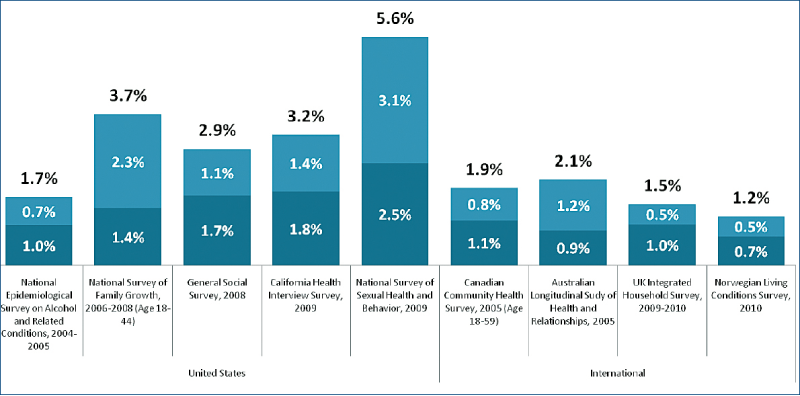
Spre deosebire de berbeci, oamenii sunt homosexuali în proporţie de 3,5% (Gates, 2011, vezi şi figura de mai jos) (incluzînd aici şi femeile lesbiene şi transgenderii), în USA sau de 4-4,1% în Franţa (Midi Libre, 3.07.2014****). O proporţie mai mare pare a exista în India: 10% din bărbaţii necăsătoriţi au întreţinut o relaţie homosexuală (Patel, 2012).

Primele studii asupra „berbecului homosexual“ au apărut încă din 1974 (Geist, citat de Roselli, 2009). Cercetările ulterioare au arătat că berbecii au, la nivelul hipotalamusului, un cluster de celule (nucleu) care este legat de sex, numit ovine SDN. La acest nivel, s-a observat că berbecii heterosexuali au o cantitate mai mare de aromatază (Roselli, 2009). Datele de la berbeci, unde dezvoltarea oSDN se face prenatal (ziua 30 – ziua 90) au fost extrapolate la om. Administrarea de testosteron la oi conduce la creşterea oSDN la fel ca la berbec (masculinizarea oSDN). Castrarea postnatală sau administrarea de testosteron postnatal nu modifică dimensiunea şi numărul de neuroni din oSDN (Roselli, 2009).
Berbecii homosexuali sunt diferiţi de cei heterosexuali, din punct de vedere hipotalamic în aria preoptică (Morris, 2004). Acest fapt a fost observat şi la şobolani şi, din nou, ulterior, la om.
La berbec (ca şi la om) se pun două probleme:
1. Orientarea (driving) sau atracţia de a face sex cu oi sau cu berbeci, în sensul de a avea comportament de berbec faţă de o oaie vs. comportament de berbec faţă de un berbec, şi nu faţă de oaie.
La berbeci, comportamentul sexual este indus hormonal. Un berbec este atras de o oaie (female driving) atunci când nivelul lui de LH şi, implicit, testosteron creşte (LH surge). Sau, altfel spus, un berbec are un raport sexual cu o oaie dacă, la nivel hipotalamic, creşte LHRH, care declanşează secreţia de LH, gonadotropul responsabil cu creşterea testosteronului. Creşterea testosteronului declanşează orientarea spre oaie a unui berbec „normal“ (Interesant este că prezenţa berbecului lângă oaie declanşează la oaie creşterea LH, care conduce la declanşarea estrului, prin creşterea nivelului de estradiol).
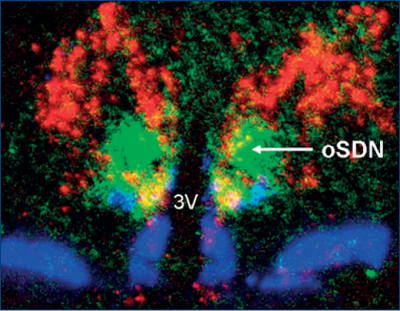

Datele obţinute de la berbecii „homosexuali“ arată că ei nu se activează sexual în prezenţa oilor la estru. Faptul sugerează fie că nu pot discrimina între feromonii estrogenci vs. cei androgenici, fie că hipotalamusul lor este insuficient sau că are nevoie de un stimul mult mai puternic pentru a se activa, i.e. orienta. Aceasta arată că berbecii orientaţi sexual spre oi au mecanisme neuro-hormonale diferite faţă de cei orientaţi spre berbeci.
Ca urmare a faptului că un berbec heterosexual caută o oaie atunci când aceasta (i.e. oaia) are estradiolul crescut, urmare a eliberării hipotalamice de LH, atunci este de aşteptat ca un berbec „homosexual“, când caută un alt berbec, o face pentru că, probabil, are loc o modificare de atracţie, cu două posibilităţi:
-
considerând că pornirea unui berbec spre o parteneră (female driving) este dependentă de creşterea testosteronului propriu, indusă de LH, ca urmare a creşterii estradiolului oii, atunci ne-am aştepta ca pornirea unui berbec „homosexual“ spre un alt berbec să fie declanşată tot de creşterea testosteronului propriu faţă de un alt fel de fenomen steroidic al unui potenţial berbec „primitor“. Acest fenomen nu a fost observat. Berbecii „homosexuali“ nu prezintă creşteri ale testosteronului la momentul „atracţiei“ către un alt berbec (de remarcat că experimentele s-au realizat în prezenţa unor oi la estru).

-
fie berbecul „homosexual“ are modificări cerebrale de miros şi este atras de testosteronul unui alt berbec, şi nu de estradiolul unei oi. Acest fenomen nu se observă însă. Un berbec „normal“ care secretă testosteron nu acceptă un berbec „homosexual“.

Aceste observaţii arată că contactul sexual la berbecii homosexuali nu este legat de activarea hipotalamusului (pe linie de releasing) şi nici de creşterea LH (hipofizar). Ipoteza a fost translatată şi investigată şi la om.
2. Acceptarea (accepting) sau fenomenul de a accepta pasiv de a face sex cu un berbec. Acest fenomen nu a fost observat la berbeci. Un berbec „normal“ care secretă testosteron nu acceptă un berbec „homosexual“. Fenomenul observat la nivelul rasei Ovis Aries nu este extrapolabil rasei Homo Sapiens, în sensul că bărbaţii homosexuali „pasivi“ (i.e. care acceptă relaţii sexuale cu un alt bărbat) au nivele de testosteron normale.

Acceptarea de a face sex cu un berbec există la specii de oi sălbatice (gen muflon, e.g. Ovis canadensis), dar fenomenul poate fi descris ca „obligaţie“ socială, şi nu sexuală; prin segregare spaţială în afara estrului oilor, berbecul alfa îi „supune“ la sex pe berbecii secundari (beta, gama) (fenomenul poate fi descris în anumite situaţii sociale umane specifice).
În schimb, experimente pe berbeci, cărora li s-a administrat estradiol, au arătat că berbecii estrogenizaţi nu acceptă nici ei berbeci „homosexuali“.
De aceea, studiile pe berbecii homosexuali nu pot decât să sugereze mecanisme, nu să şi aducă dovezile supreme. La om, comportamentul sexual şi preferinţa pentru un anumit tip de partener sunt disociate şi par a fi reglate de alte mecanisme.
De aceea, în ceea ce priveşte orientarea sexuală, şi, de aici, şi tulburarea de orientare sexuală, teoriile strict hormonale au fost combătute de teorii ale comportamentului, ce ţin de sfera cercetării psihiatrice, de dezvoltarea şi achiziţia socială, ca de exemplu teoria controlului bio-psiho-social a lui Friedman (1993).
Cu toate acestea, adepţii teoriilor pur biologice afirmă că nu există date certe care să probeze că mecanismele de orientare sexuală ar fi de origine postpartum (i.e, factori de mediu, factori sociali; Swaab, 2002, Savic, 2010), chiar dacă diferenţierea şi maturarea sexuală a hipotalamusului se petrece între vîrsta de 4 ani şi ajungerea la maturitate (Swaab, 2002), deci mult mai tîrziu decât se presupunea. Dacă fatorii postnatali acţionează, ei sunt numai factori favorizanţi, şi nu determinanţi.
Datele de observaţie ştiinţifică acumulate în ultimii ani susţin mai degrabă teoriile „hormonale“ asupra homosexualităţii, sugerând că homosexualii prezintă modificări de tip feminin ale dezvoltării cerebrale şi că prezintă un dimorfism specific. Însă, prin termenul de „teorie hormonală“ trebuie înţeles tot ceea ce ţine de „unitatea operaţională hormon – receptor“, definită ca atare încă din 1992 (Grigorescu, 1992).
Argumente ale teoriei hormonale
a homosexualităţii
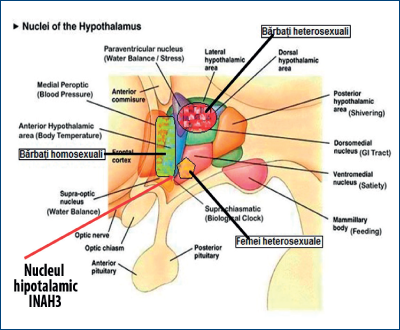
Argumentul 1. Primele cercetări clarificatoare asupra dimorfismului neurocomportamental au fost iniţiate în 1985 (vezi în Morris, 2004) şi au arătat că al treilea nucleu interstiţial al hipotalamusului anterior (INAH3) este mult mai mare la bărbat decât la femeie. Acest nucleu a fost considerat a fi corespondentul oSDN de la berbec.
Argumentul 2. În 1991, Simon LeVay (vezi în Morris, 2004) nu numai că a replicat cerecetările anterioare, ci a arătat că acest nucleu este, la homosexualii bărbaţi, identic cu cel de la femei. Aceasta a sugerat că persoanele cu un nucleu INAH3 mare sunt atrase de femei, iar cele cu INAH3 mic sunt atrase de bărbaţi. În acest nucleu s-au observat multe celule/neuroni producătoare de hormon antidiuretic (vasopresină).
Argumentul 3. Apoi, s-a pus problema dacă acest fenomen este consituţional sau este indus de homosexualitate. S-a arătat că prezenţa de mai multe celule (neuroni) producătoare de hormon antidiuretic (ADH) (vasopresină) în hipotalamusul homosexualilor vs. heterosexuali nu este un fenomen indus de homosexualitate, ci un predat, antenatal (Swaab, 2008). Importanţa serică a ADH în exprimarea sexului nu a fost până acum abordată (cercetarea pe www.pubmed.gov la „siadh, sex“ sau „desmopresin, excess, sex“ nu are nicio valoare), deşi prevalenţa crescută în rândul femeilor a SIADH a fost citată (Cumming, 2014).
Motivul pentru care aceşti nuclei capătă o dezvoltare aşa de mare la bărbaţi este determinat de un moment limitat în timp, în viaţa intrauterină. Atunci, o anumită şi adecvată cantitate de androgen(i) irigă acea zonă, androgeni depenenţi de activitatea intrauterină a gonadei (testiculului). Cel puţin pentru berbeci, lucrurile sunt acum lămurite, chiar dacă problema a făcut valuri sociale de la dreapta la stânga şi în disputele dintre liberali şi tradiţionalişti (Cîmpeanu, 2007).
Argumentul 4. Implicarea hormonilor postpartum a fost înlăturată pentru că nu s-a citat o frecvenţă mai mare a homosexualităţii printre subiecţii care au avut „criză estrogenică“ la naştere (ginecomastie postpartum), deşi se ştie că aceasta apare la 70% din nou-născuţi (Amer, 2009) şi nici nu se ştie ca eunucii să fie mai homosexuali decât bărbaţii cu testiculele în locul lor (Aucoin, 2006).
Privind spre normalul biologic, s-a propus o dublă curbă Gauss-Laplace, în care „centrul“ este nivelul de testosteron necesar pentru a produce „androgenizarea“ creierului (figura 22).
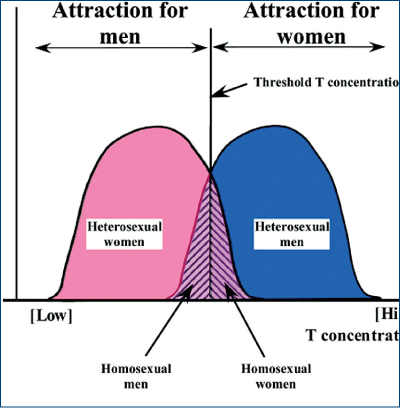
Cu toate acestea, cercetătorii nu sunt încă în consens cu privire la om, pentru că evidenţe arată că procesul de formare a nucleului hipotalamic INAH3 nu ţine neapărat de nivelul de androgeni: homosexuali pot apărea şi prin expunere la puţini androgeni, dar şi la mulţi androgeni.
Argumentul 5. Date anatomopatologice efectuate postmortem pe diferiţi subiecţi arată că INAH3 (corespondentul oSDN) este mai mic ca dimensiune la homosexualii cu AIDS decedaţi (nr. 14) faţă de heterosexualii cu aceeaşi boală (nr. 10) şi faţă de heterosexualii fără AIDS (nr. 24) (Byne, 2001). Cu toate acestea, numărul de neuroni per frotiu nu a fost diferit în cazurile analizate, sugerând că, dacă o leziune este prezentă, ea se manifestă calitativ, şi nu cantitativ.
Argumentul 6. Date care să ateste hiperandrogenizarea femeilor devenite lesbiene (de tipul raportul index-inelar, emisii oto-acustice, modificate în sens de masculinizare) nu pot fi reproduse în studii multiple (vezi în Balthazart, 2011).
De aceea, teoria hormonilor circulanţi nu poate susţine singură cauza homosexualităţii masculine. Alta trebuie să fie explicaţia!
Argumentul 7. Într-adevăr, cercetările lui Resko (1996) şi Roselli (2004, vezi mai departe şi în Morris, 2004) au arătat (la berbec) că nu androgenii în sine, adică cei circulanţi, sunt responsabili de dezvoltarea comportamentului promasculin, ci nivelul de aromatază, enzima care converteşte testosteronul la estradiol!
Cu toate acestea, nivelul de conversie de la testosteron spre estradiol, la nivel cerebral, pare a depinde totuşi şi de nivelul de androgeni circulanţi (Resko, 1996). Oricum, nivelul de aromatază este dependent de cromozomul X, ceea ce sugerează că homosexualii (berbeci) apar aşa prin transmisie maternă (precum este sugerat şi la om, vezi mai departe)! Încă o dată, este mai greu să fii bărbat decât femeie.
De aici, ipoteza că homosexualii reprezintă un continuum de dezvoltare de la femeie la bărbat.
Cel mai probabil este că şi la om există astfel de modificări. Dar, în acest caz, diagnosticul înseamnă a preleva creierul, a-l îngloba în parafină şi a cerceta imunocitochimic prezenţa şi cantitatea de aromatază, ceea ce este evident că nu poate fi vorba de diagnostic, ci numai ca studiu retrospectiv după moarte (Kruijver, 2001).
Primele cercetări, care au implicat hormonii din sânge, nu au arătat diferenţe biologice între grupurile comportamentale. Chiar şi nivelul de receptori în diferite arii hipotalamice nu arată diferenţe de orientare sexuală, ci numai de nivel hormonal (Kruijver, 2001).
În ultimii ani, însă, tehnologii noi de imagistică cerebrală au permis observarea reacţiilor diferitelor arii cerebrale şi hipotalamice (figura 23), precum şi studierea nivelurilor locale de neurotransmiţători în condiţiile diferiţilor stimuli, sexuali sau indiferenţi.
Aceste cercetări au arătat diferenţe clare între bărbaţii hetereosexuali şi cu orientare bărbătească clară, faţă de homosexuali şi transsexuali.
Argumentul 8. De exemplu, nucleul suprachiasmatic şi comisura anterioară sunt mai mari la homosexuali decât la heterosexuali (Kinnunen, 2004).
Interesant este că numai anumite arii cerebrale prezintă diferenţe. De exemplu, activitatea hipotalamusului pe linie de stimulare a corticosuprarenalei (prin corticotropin-releasing hormon, CRH) şi tiroidiei (thyroxine-releasing hormon, TRH) este identică şi la heterosexuali şi la transsexuali. Aceşti hormoni nu sunt implicaţi în formarea sau suţinerea normalităţii sexului neurocomportamental (indiferent activity).
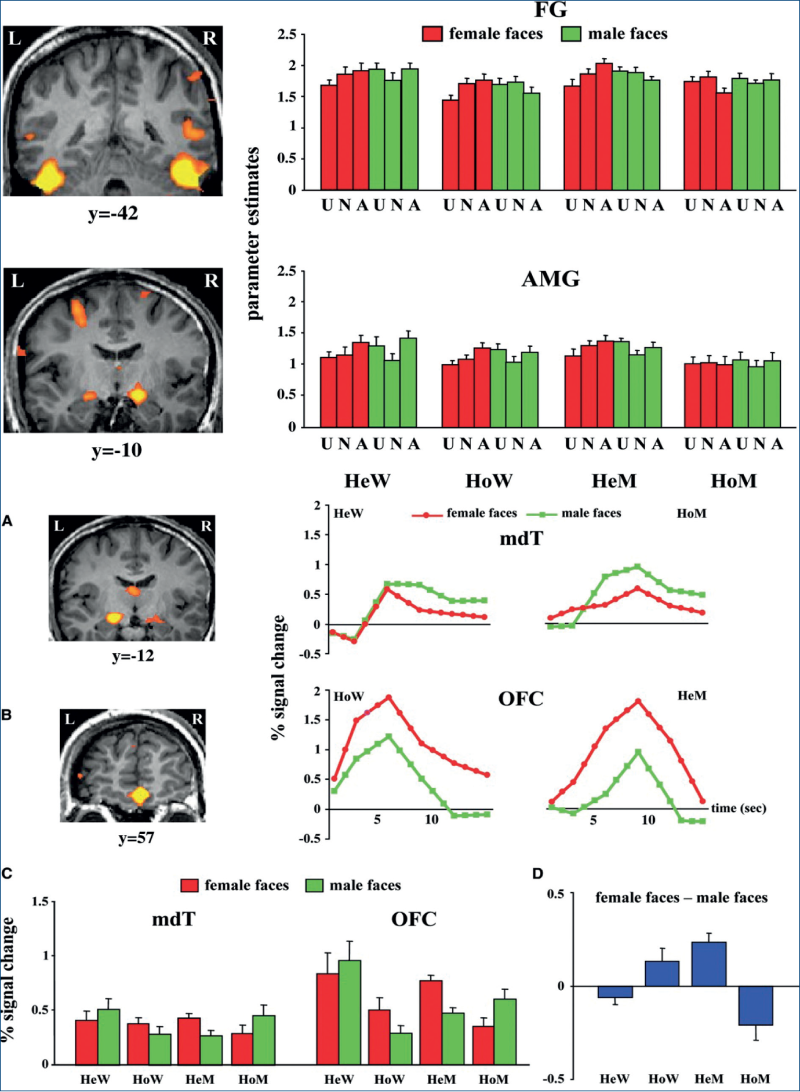
Argumentul 9. Cea mai diferită arie hipotalalmică activă investigată în vederea stabilirii diferenţelor de comportament sexual, prin utilizarea de feromoni, folosind imagistica funcţională cu positron emission tomography (PET), a fost observată ca fiind aria medială preoptică a hipotalamusului anterior (figura 8) (Savic, 2005). Efectele feromonilor „sexuali“ pot fi sintetizate astfel:
-
femeile au fost supuse la miros de feromoni cu:
-
androgeni (4,16-androstadien-3-one); efectul acestora a fost de a activa zone active la heterosexuale şi lesbiene;
-
estrogeni (1,3,5(10),16-tetraen-3-ol); efectele acestui estrogen au fost neconcludente la lesbiene şi negative la heterosexuale;
-
bărbaţii au fost supuşi la miros de feromoni cu:
-
estrogeni (1,3,5(10),16-tetraen-3-ol); aceşti estrogeni au generat zone active numai la heterosexuali adevaraţi;
-
androgeni (4,16-androstadien-3-one); androgenii au activat zonele hipotalamice numai la homosexuali pasivi.
De aici rezultă clar că există diferenţe în exprimarea neuroendocrină a bărbaţilor heterosexuali faţă de cei homosexuali pasivi. De observat că mirosuri curente sau chiar parfumuri comerciale, considerate „atractive“, nu au efecte diferite asupra ariilor hipotalamice investigate (Savic, 2005); în plus, s-a dovedit că toate aceste mirosuri, inclusiv feromonii, activează numai ariile olfactive: amigdala, aria piriformă, aria orbito-frontală şi cortexul insular.
Argumentul 10. Investigaţia vizuală cu RMN funcţional cerebral (figura 24) arată date similare: arii cerebrale (nucleul mediodorsal al talamusului şi cortexul medial orbitofrontal) de la heterosexualii bărbaţi şi homosexualele femei se activează la văzul unei femei, în timp ce aceleaşi arii cerebrale la homosexualii bărbaţi şi heterosexualele femei se activează la văzul unor fotografii de bărbaţi (Kranz, 2006).
Argumentul 11. Alte observaţii prin PET arată că heterosexualii metabolizează diferit fluoxetina (un SSRI) faţă de homosexuali, fenomen observabil cu PET cu fluorodeoxiglucoză (Kinnunen, 2004).
Legat de comportamentul sexual, o cercetare interesantă, legată şi de grupul suedez (Savic), este cea care a arătat că, prin miros, o şoricioaică poate să avorteze dacă în preajma ei nu se află tatăl copiilor ei, ci un alt mascul (Hatt, 2007). Concluzia filozofică a fost că bărbatul trebuie să-şi protejeze familia!
De aici, necesitatea de a cerceta mult mai multe arii cerebrale, căci şi ele pot fi implicate în orientarea sexuală. Şi de asemeni important este de a cerceta larg procesele de recepţie, aromatizare şi hidrogenare a testosteronului de la nivelul acestor arii.
Argumentul 12. În plus, alţi neurohormoni în afara celor „sexuali“ pot fi implicaţi în comportamentele sexuale homosexuale. De exemplu, scăderea nivelului celular al dopaminei la Drosophila conduce la generarea de masculi cu orientare homosexuală (Liu, 2009), în sensul creşterii atracţiei pentru alţi masculi ce nu au fost modificaţi (wild type) şi de scădere a adversităţii faţă de masculii orientaţi spre masculi. Modificarea indusă prin scăderea dopaminei nu modifică din atracţia animalelor modificate pentru animalele modificate sau nemodificate. Fenomenul sugerează că un comportament normal sexual de tip masculin poate fi canalizat spre sexul masculin dacă animalul de experienţă este modificat adecvat (Liu, 2009).
În concluzie, se poate admite că homosexualitatea, cel puţin masculină, este un tip de pseudohermafroditism şi reprezintă o patologie a sexului neurocomportamental.
Cu toate acestea, fenomenul poate fi privit ca pozitiv. Se ştie că femeile sunt mai deştepte decât bărbaţii, că au o capacitate de cogniţie mai mare, generată de nivelul de estrogeni*****, şi că mari realizatori artistici, bărbaţi homosexuali, au reuşit pe acest câmp de exprimare umană tocmai datorită organizării feminine a creierului.
O temă interesantă în dezbatere este cea care sugerează că homosexualii bărbaţi (pasivi) ar fi predispuşi către artă şi că sunt importanţi creatori artistici. Printre argumente stau „nume“ grele din pictură (Leonardo da Vinci, Caravaggio), sculptură (Michelangelo) etc. Printre argumentele pentru susţinerea unor astfel de afirmaţii sunt cele care ţin de modul de redare al portretisticii; pictori (bărbaţi) care pictează nuduri de bărbaţi/băieţi sau bărbaţi cu aspecte feminine, sau portrete de femei considerate a fi de bărbaţi (vezi disputa asupra identităţii Giocondei) (Blum, 2001).


La astfel de argumente se poate replica cu antonimul: numărul de pictori bărbaţi care pictează nuduri de femei. Nu ştiu să existe o analiză statistică pe această temă.
Un alt gen de argument ar fi că homosexualii sunt mai mult stângaci (circa 14%) şi că stângacii sunt mai creativi (Götestam, 2001); dar aceiaşi homosexuali sunt şi mai predispuşi la gângăvit (stuttering) şi la dislexie (Götestam, 2001), precum şi la ciroză biliară, boală Crohn şi rectocolită ulcerohemoragică (Searleman, 1987).
Pe de altă parte, nu există date ştiinţifice care să ateste faptul că artiştii ar fi mai mult homosexuali decât heterosexuali (Demb, 1992). Un argument statistic de nerefuzat ar fi acela dacă numărul de artişti homosexuali depăşeşte cu mult estimarea de circa 4% pentru homosexualitate în populaţia generală. Printr-un test z (de analiză a procentelor) sau test X2 (de analiză a numerelor pe 2 coloane şi 2 rânduri), acest fapt poate fi dovedit. Nu cunosc ca în acest moment să se fi făcut o astfel de cercetare.
Cauze biologice ale homosexualităţii
1. Condiţionări genetice. În afara cercetărilor cu impact direct pe modelul dezvoltării orientării sexuale, s-a observat, la modul general, că comportmentul homosexual este condiţionat genetic pe linie maternă: bărbaţii homosexuali au mai mulţi homosexuali printre rudele materne (veri, unchi) decât pe linie paternă (detalii în Balthazart, 2011). Se pare că gene situate pe diferiţi cromozomi, precum 7q36, 8p12 şi 10q26, Xq28 ar fi responsabile de aceste comportamente (Rodríguez-Larralde, 2009).
2. Cauze imune. Se ştie şi că într-o fratrie multiplă, fraţii mai mici au mai mari şanse să devină homosexuali faţă de fraţii mai mari, fenomen cunoscut ca fraternal birth order effect (Blanchard, 2014). S-a sugerat că fenomenul ţine de blocarea prin anticorpi antisex masculin a gonadei (antiH-Y), anticorpi ce apar la mamă în sarcinile multiple cu băieţi (Blanchard, 2001).
Ipoteza leziunii imune materne poate explica constatarea că bărbaţii heterosexuali cu fraţi mai mari cântăresc la naştere mai puţin decât bărbaţii heterosexuali cu surori mai mari şi că bărbaţii homosexuali cu fraţi mai mari cântăresc chiar mai puţin decât bărbaţii heterosexuali cu fraţi mai mari (Blanchard, 2001, 2014).
Sugestii recente au arătat că nu neapărat nivelul scăzut de testosteron poate fi cauza alterării circuitelor cerebrale şi hipotalamice în sens homosexual, ci leziuni directe la nivelul sistemului nervos (probabil prin autoanticorpi specifici, blocaje de receptori androgenici) (Bogaert, 2011).
Ca antinomie faţă de cercetările de patogenie biologică a homosexualităţii s-au încercat explicaţii psihologice. Printre ele, s-a sugerat că abuzul sexual în copilărie ar putea fi o cauză, atât pentru homosexualitate, cât şi pentru pedofilie (James, 2006). Cu toate acestea, s-a arătat că majoritatea semnificativă a băieţilor abuzaţi în copilărie nu devin homosexuali (James, 2006).
B. Feminin
Cercetările homosexualităţii feminine sunt mai puţine decât cele ale homosexualităţii masculine. Se pare că numărul de lesbiene este mai mic decât cel al bărbaţilor homosexuali, sugerând că pattern‑ul feminin este chiar feminin şi mai greu de controlat hormonal sau social.
Multe din cazurile de femei cu homosexualitate activă au fost citate în anumite situaţii clinice specifice. Două astfel de situaţii au fost observate: hiperplazia de corticosuprarenală, administrarea la mamă de dietilstilbestrol, la care se poate adăuga non-lesbianismul din sindromul Turner.
1. S-a arătat că femeile (paciente cu sex cromozomial 44XX) cu hiperplazie de corticosuprarenală congenitală (prin deficit de 21-hidroxilază) se comportă mult mai frecvent ca lesbiene proactive de tip masculin decât surorile lor neafectate de această boală virilizantă (Hines, 2006). Proporţia de homosexualitate pură (100% din raporturile seuxale) este de 6,25%, homosexualitatea mai mare de 75% este de 12,5% şi tot 12,5% dintre aceste femei sunt 50% homosexuale/heterosexuale. Interesant este că numai 5% din femeile cu hiperplazie de corticosuprarenală congenitală doresc să-şi schimbe sexul.
2. În anii ’60, pentru preîntâmpinarea avortului în sarcinile cu problemă s-a administrat dietilstilbestrol (DES). Fetele născute din mame cărora li s-a administrat DES s-au arătat a fi mult mai frecvent lesbiene decât semenele lor (vezi în Balthazart, 2011). Fenomenul nu a fost pe deplin explicat, pentru că s-a considerat că acţiunea de masculinizare a creierului este dependentă de aromatizarea de novo a estradiolului intracerebral, şi nu de niveluri crescute de estrogeni în sânge.
3. Ca argument pentru condiţionarea genetică şi prenatală a sexului este faptul că lesbianismul nu a fost citat în sindromul Turner pur (persoane genetic X0), în care fenotipul este feminin, dar şi comportamentul sexual este de tip feminim. Atunci când, la un caz de sindrom Turner, s-a observat comportament homosexual (lesbian), cercetările mai aprofundate au arătat că „femeia“ prezenta mozaicism, adică prezenta gene de tip masculin provenite de la un cromozom Y, ataşate de somatozomi (Fishbain, 1980).
Cercetările de imagistică cerebrală funcţională cu stimulente sexuale au adus date noi privind diferenţa între femeile heterosexuale şi cele homosexuale (lesbiene).
Spre deosebire de bărbaţii homosexuali, care arată modificări foarte clare în activarea ariilor optice hipotalamice (Savic, 2005), femeile lesbiene nu prezintă astfel de dimorfisme; ele acţionează ca femei, atunci când sunt supuse la estrogeni (Berglund, 2006). Adică, după mirosul estrogenilor, atât femeile heterosexuale, cât şi lesbienele nu sunt excitate hipotalamic. Fenomenul este neobişnuit, căci suprapunerea cu fenomenul de la bărbaţi ar fi trebuit să conducă la acest comportament.
În schimb, lesbienele diferă de femeile heterosexuale prin faptul că nu reacţionează prin aria optică hipotalamică la androgeni. De fapt, ele reacţionează la androgeni, dar nu prin aria considerată ca normală pentru un asemenea comportament sexual (Berglund, 2006).
Femeile lesbiene se activează la nivelul hipotalamusului dorsomedial şi paraventricular (VOI) cu estrogeni, la fel ca şi bărbaţii heterosexuali, dar diferit de femeile heterosexuale. Toate aceste date sugerează că femeile lesbiene procesează atât androgenii, cât şi estrogenii mai asemănător cu bărbaţii heterosexuali decât cu femeile heterosexuale (Berglund, 2006).
Femeile homosexuale prezintă arii cerebrale de substanţă cenuşie precum bărbaţii heterosexuali (Ponseti, 2007, vezi figurile de mai jos): cortexul temporo-bazal, zona ventrală a cerebelului şi cortexul ventral sunt premotor.
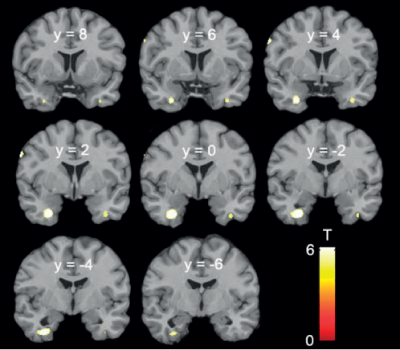
Faptul sugerează că pentru un comportament de tip bărbătesc este necesar ca acele arii să fie astfel construite. Implicaţia ar fi că, dacă testosteronul acţionează la bărbaţii heterosexuali pentru a determina comportament heterosexual, atunci acele arii apar aşa tot prin acţiunea testosteronului endogen la fătul de sex feminin.
Cu toate acestea, bărbaţii homosexuali nu arată modificări în acele arii, la fel cum nu arată nici bărbaţii heterosexuali. Aceasta arată că acele arii au fost testosteronizate prenatal la bărbaţii homosexuali. Aceasta arată şi că mecanismele prin care apare homosexualitatea masculină sunt diferite de cele prin care apare homosexualitatea feminină sau că fenomenul poate fi influenţat de niveluri diferite de androgen prenatal.
V. Tulburări ale identităţii de sex/gen
Cercetările asupra identităţii de sex nu sunt decât umane. Nu există model animal, pentru că, până acum, nici un animal nu a cerut schimbarea de sex!
Tulburarea de identitate de sex (mai corect ar fi „de gen“) sau transsexualismul se referă la acei pacienţi/acele paciente care, în pofida dezvoltării sexuale aparent normale şi chiar a unei vieţi sociale normale (căsătoriţi, cu copii), cer să aibă celălalt sex, pentru că nu se identifică cu sexul biologic sau cu cel asumat social. Există un sinonim creat de psihiatri: disforie sexuală.
Prevalenţa a fost estimată la 0,00341% în Irlanda (apreciată ca „normală“ pentru Europa) (Judge, 2014).
Cauzele pentru care apar astfel de pacienţi au fost sugerate a fi psihologice, în context social şi cultural, şi biologice. Din punct de vedere biologic, cel mai implicat factor care pare a fi responsabil de tulburările de identificare sexuală, de gen, ţine de faptul că organele genitale se diferenţiază în lunile 1-3, în timp ce programarea sexuală a creierului apare mai târziu, în lunile 4-6. De aici faptul că gradul de masculinizare al organelor genitale (sexul gonadal şi cel al organelor genitale) poate să nu reflecte gradul de masculinizare al creierului (Savic, 2010). În plus, formarea penisului este sub controlul unei hidrogenenaze (5 alfa-reductază), în timp ce creierul este sub controlul unei aromataze (vezi şi mai sus şi figura de mai jos) care transformă Testo în E2.

Se afirmă că nu există nici o dovadă că mediul postpartum (fie el mai precoce sau tardiv) are efect asupra identităţii sexuale, dar nici asupra orientării sexuale. Până la ce nivel al observaţiei este necesar a ajunge pentru a dovedi orientarea programată a sexului este încă o temă de discuţie. Cercetări recente arată din ce în ce mai mult dimorfisme cerebrale tipice, dar şi atipice (la subiecţi homosexuali şi transgenderi). Care sunt circuitele neuronale responsabile pentru aceasta este încă neclar (Savic, 2010). Printre candidaţi figurează amigdala, girusul cingular anterior, putamenul, caudatul, cortexul frontal (Savic, 2008).
A. Masculin
Transsexualismul masculin este mult mai frecvent. Raportul cu transsexualismul feminin este de 3-2:1 (Tugnet, 2007, Judge, 2014).
Pentru a găsi cauze biologice la acest fenomen clinic, au fost folosite mai multe modele de cercetare.
Un model a fost legat de nucleul supracapsular al stria terminalis (BSTc), unde au fost studiate numărul de celule şi cantitatea de somatostatină conţinută (vezi în Swaab, 2004). Astfel, s-a arătat că numărul neuronilor din acest nucleu creşte cu vârsta. Volumul acestui nucleu este dublu la bărbaţi faţă de femei şi conţine o cantitate dublă de somatostatină. La transsexualii bărbaţi ce se simt ca femei (M∞F), şi care cer a deveni femei, acest raport este inversat, aşa cum este la femeile normale.
Interesant este că, din punctul de vedere al modelului BSTc, bărbaţii heterosexuali nu diferă de bărbaţii homosexuali, care se cred în continuare bărbaţi!
Un alt teritoriu implicat în tulburările de identificare a sexului este corpul calos. Pacienţii cu tulburări de identificare de sex care sunt genetic bărbaţi, dar care se identifică femei, prezintă o funcţionalitate a corpului calos precum femeile, iar pacientele care sunt genetic femei, dar care se identifică ca bărbaţi, prezintă o funcţionalitate a corpului calos precum bărbaţii. Or, se ştie că corpul calos este un important sediu al dimorfismului sexual bărbat-femeie, fiind locul prin care se transmit informaţiile de la cele două emisfere, la rândul lor organizate asimetric la bărbat faţă de femeie (vezi în Swaab, 2008). Mecanismele prin care se produce acest fenomen nu sunt încă pe deplin elucidate (Yakota, 2005).
Prin RMN funcţional s-a arătat că transsexualii au un grad mai mare de centralizare la nivelul lobului parietal superior.
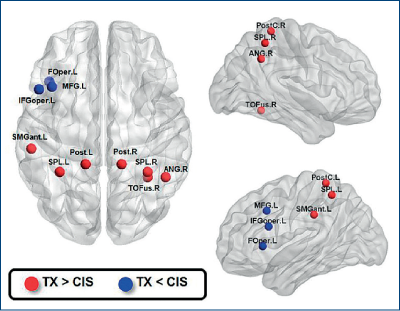
Conectivitatea dintre insula dreaptă şi cortexul primar somatosenzitiv a fost negativ corelată cu aprecierea sexului/genului dorit. Datele arată că există un grad de disociere între corp şi identitate (Lin, 2014, figura 31).
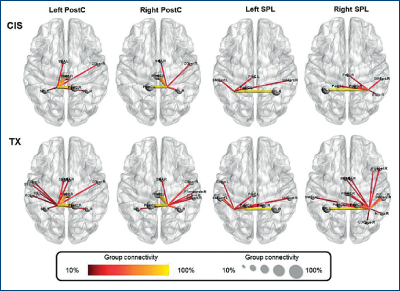
B. Feminin
Tulburarea de gen feminin este mai rară. Mai puţine femei/fete doresc să devină bărbaţi. Datele observaţionale arată că fenomenul este mai mult biologic decât psihologic.
Dovezile clinice sunt concordante cu teoria că sexul este definit antepartum. Sexul neurocomportamental masculin se poate manifesta la femei, dacă femeia a fost impregnată testosteronic antepartum, intrauterin.
În ceea ce priveşte tulburarea de identitate de sex feminin, trei situaţii clinico-biologice au fost până acum identificate.
1. De mult se ştie că 50% din „fetele“ care cer să devină băieţi sunt de fapt organisme XY (detalii în Balthazart, 2011). Cel mai nou caz este al lui Mark şi Pam Crawford, citat în documentele Eur. Soc. Endocrinol. (figura 33) care au înfiat şi crescut „o fetiţă“ de la 2 ani, pentru ca la vârsta de 7 ani „aceasta“ să ceară să fie „băiat“. Atunci s-a aflat că era de fapt un organism XY cu un fost testicul extras imediat după naştere, cu disgenezie genitală (vezi figura 32), organism crescut ca fată.

Întrebarea pertinentă este: dar de ce nu toate persoanele XY, crescute ca fete cu disgenezii genitale recorectate de la băiat la fată, nu cer modificarea sexului spre cel bărbătesc? Răspunsul se aşteaptă a fi dat în viitor.
2. Cazuri mai rare de transsexualism feminin au fost citate în autism (Lemaire, 2014). Cu toate acestea, se ştie că autismul este mult mai frecvent la băieţi şi că aceştia au şi un comportament masculin mai agresiv, sugerând un rol pentru testosteronul intrauterin la femeile cu tulburarea de sex.
3. La transsexualii femei care se simt bărbaţi şi cer să fie considerate bărbaţi (F ➞ M), volumul nucleului supracapsular al stria terminalis (BSTc) este identic cu cel al bărbaţilor heterosexuali normali (Swaab, 2004). n
Bibliografie
- Alexander GM. Postnatal testosterone concentrations and male social development. Front Endocrinol (Lausanne), 2014, Feb 21; 5, 15: 1-6.
- Amer A, Fischer H. Images in clinical medicine. Neonatal breast enlargement. N Engl J Med., 2009, Apr 2; 360, 14: 1445.
- Aucoin MW, Wassersug RJ. The sexuality and social performance of androgen-deprived (castrated) men throughout history: implications for modern day cancer patients. Soc Sci Med., 2006, Dec; 63, 12: 3162-3173.
- Balthazart J. Minireview: Hormones and human sexual orientation. Endocrinology, 2011, Aug; 152, 8: 2937–2947.
- Berenbaum SA, Beltz AM. Sexual differentiation of human behavior: effects of prenatal and pubertal organizational hormones. Front Neuroendocrinol., 2011, Apr; 32, 2: 183-200.
- Berglund H, Lindstrom P, Savic I. Brain response to putative pheromones in lesbian women. ProcNatAcadSsci USA, 2006, May 23, 103, 21: 8269–8274.
- Blanchard R. Fraternal birth order and the maternal immune hypothesis of male homosexuality. Horm Behav., 2001, Sep; 40, 2: 105-114.
- Blanchard R. Detecting and correcting for family size differences in the study of sexual orientation and fraternal birth order. Arch Sex Behav., 2014, Jul; 43, 5: 845-852.
- Bogaert AF1, Skorska M. Sexual orientation, fraternal birth order, and the maternal immune hypothesis: a review. Front Neuroendocrinol., 2011, Apr; 32, 2: 247-254.
- Blum HP. Pychoanalysis and art, Freud and Leonardo. J Am Psychoanal Assoc., 2001, Fall; 49, 4: 1409-1425.
- Byne W, Tobet S, Mattiace LA, Lasco MS, Kemether E, Edgar MA, Morgello S, Buchsbaum MS, Jones LB. The interstitial nuclei of the human anterior hypothalamus: an investigation of variation with sex, sexual orientation, and HIV status. Horm. Behav., 2001, Sept; 40, 2: 86-92.
- Cîmpeanu C. Berbecul homosexual încinge America. România liberă (Buc.), Marti, 6 Februarie 2007.
- Cornil CA, Ball GF, Balthazart J, Charlier TD. Organizing effects of sex steroids on brain aromatase activity in quail. PLoS One., 2011, Apr 29; 6, 4: e19196.
- Cumming K, Hoyle GE, Hutchison JD, Soiza RL. Prevalence, incidence and etiology of hyponatremia in elderly patients with fragility fractures. PLoS One, 2014, Feb 5; 9, 2: e88272.
- Demb J. Are gay men artistic? A review of the literature. J Homosex., 1992; 23, 4: 83-92.
- Den Hond E, Schoeters G. Endocrine disrupters and human puberty. Int J Androl. 2006 Feb;29(1):264-71.
- Fishbain DA, Vilasuso A. Exclusive adult lesbianism associated with Turner’s syndrome mosaicism. Arch Sex Behav., 1980, Aug; 9, 4: 349-353.
- Friedman RC, Downey J. Neurobiology and sexual orientation: current relationships. J Neuropsychiatry Clin Neurosci., 1993, Spring; 5, 2: 31-53.
- Gates GJ. How many people are lesbian, gay, bisexual, and transgender The Williams Institute, UCLA School of Law, Los Angeles, CA, USA. williamsinstitute@law.ucla.edu, www.law.ucla.edu/williamsinstitute, 2011.
- Geist, V. Mountain Sheep A Study in Behavior and Evolution. The University of Chicago Press; Chicago: 1974. p. 139-2554.
- Georgescu C. Endocrinopatiile şi disfunctiile cuplului sexual. A V-a Conf.Naţ. AMSR, Sinaia, 28.10.2005.
- Gooren L. Is there a hormonal basis for human homosexuality? Asian J Androl., 2011, Nov; 13, 6: 793-794.
- Götestam KO. Handedness and creativity in a sample of homosexual men. Percept Mot Skills, 2001, Jun; 92, 3 Pt 2: 1069-1074.
- Grigorescu A., Pereţianu D. Organizarea generală a sistemului endocrin. Celula endocrină. Celula ţintă hormonal-receptoare. Unitatea operaţională hormon-receptor. În Tratat de Endocrinologie Clinică (Treatise of Clinical Endocrinology) S.M.Milcu (ed.) I, Editura Academiei, Bucureşti, 1992, pp 29-89.
- Hatt H. Odorant receptors and reproduction. Endocrine Abstracts, 2007, 14, S.21.1.
- He FQ, Zhang HR. Perinatal clomiphene citrate treatment changes sexual orientations of male mice. Dongwuxue Yanjiu. 2013 Oct;34(5):487-92.
- Hines M. Prenatal testosterone and gender-related behaviour. Eur J Endocrinol., 2006, Nov; 155, Suppl 1: S115-S121.
- James WH. Two hypotheses on the causes of male homosexuality and paedophilia. J Biosoc Sci., 2006, Nov; 38, 6: 745-761.
- Judge C, O’Donovan C, Callaghan G, Gaoatswe G, O’Shea D. Gender dysphoria - prevalence and co-morbidities in an irish adult population. Front Endocrinol. (Lausanne), 2014, Jun 13; 5: 87-98.
- Keefe DL. Sex hormones and neural mechanisms. Arch Sex Behav., 2002, Oct; 31, 5: 401-403.
- Kinnunen LH, Moltz H, Metz J, Cooper M. Differential brain activation in exclusively homosexual and heterosexual men produced by the selective serotonin reuptake inhibitor, fluoxetine. Brain Res., 2004, Oct 22; 1024, 1-2: 251-4.
- Kranz F, Ishai A. Face perception is modulated by sexual preference. Curr Biol., 2006, Jan 6; 16, 1: 63–68.
- Kruijver FPM, Fernández-Guasti A, Fodor M, Kraan EM, Swaab DF. Sex Differences in androgen receptors of the human mamillary bodies are related to endocrine status rather than to sexual orientation or transsexuality. J.Clin.Endocrinol.Metab., 2001, 86, 2: 818-827.
- Lemaire M, Thomazeau B, Bonnet-Brilhault F. Gender identity disorder and autism spectrum disorder in a 23-year-old female. Arch Sex Behav., 2014, Feb; 43, 2: 395-398.
- Lin CS, Ku HL, Chao HT, Tu PC, Li CT, Cheng CM, Su TP, Lee YC, Hsieh JC. Neural network of body representation differs between transsexuals and cissexuals. PLoS One, 2014, Jan 20; 9, 1: e85914.
- Liu T, Dartevelle L, Yuan C, Wei H, Wang Y, Ferveur JF, Guo A. Reduction of dopamine level enhances the attractiveness of male Drosophila to other males. PLoS One, 2009, 4, 2: e4574.
- Manhães AC, Medina AE, Schmidt SL. Sex differences in the incidence of total callosal agenesis in BALB/cCF mice. Neurosci Lett., 2002, Jun 14; 325, 3: 159-162.
- Mogoş V. Androgenii. Al IV-lea Simp.Naţ. AMSR (ARDE), Sinaia, 12-13.11.2004.
- Morris JA, Gobrogge KL, Jordan CL, Breedlove SM. Brain aromatase: dyed-in-the-wool homosexuality. Endocrinology, 2004, 145, 2: 475–477.
- Patel VV, Mayer KH, Makadon HJ. Men who have sex with men in India: a diverse population in need of medical attention. Indian J Med Res., 2012, Oct; 136, 4: 563-70.
- Pereţianu D. Fiziopatologia integrării endocrine. În Tratat de Fiziopatologie (Treatise of Pathophysiology), M. Saragea (ed.), I, Editura Academiei, Bucureşti, 1985, 645-820.
- Ponseti J, Siebner HR, Klöppel S, Wolff S, Granert O, Jansen O, Mehdorn HM, Bosinski HA. Homosexual women have less grey matter in perirhinal cortex than heterosexual women. PLoS One, 2007, Aug 22; 2, 8: e762.
- Resko JA, Perkins A, Roselli CE, Fitzgerald JA, Choate JVA, Stormshak F. Endocrine correlates of partner preference behavior in rams. Biol.Reprod., 1996, 55, 120-126.
- Invest Clin. 2009 Sep;50(3):377-91. Influence of genetic factors on human sexual orientation. Review. Article in Spanish
- Rodríguez-Larralde A1, Paradisi I, Roselli CE, Larkin K, Schrunk JM, Stormshak F. Sexual partner preference, hypothalamic morphology and aromatase in rams. Physiol Behav., 2004, Nov 15; 83, 2 :233-45.
- Roselli CE, Stormshak F. The neurobiology of sexual partner preferences in rams. Horm Behav., 2009, May; 55, 5: 611-620.
- Savic I, Berglund H, Lindstrom P. Brain response to putative pheromones in homosexual men. ProcNatAcadSsci USA, 2005, 102, 20: 7356–7361.
- Savic I, Lindstrom P. PET and MRI show differences in cerebral asymmetry and functional connectivity between homo- and heterosexual subjects. Proc Natl Acad Sci USA, 2008, Jul 8; 105, 27: 9403–9408.
- Savic I, Garcia-Falgueras A, Swaab DF. Sexual differentiation of the human brain in relation to gender identity and sexual orientation. Prog Brain Res., 2010, 186: 41-62.
- Searleman A, Fugagli AK. Suspected autoimmune disorders and left-handedness: evidence from individuals with diabetes. Crohn’s disease and ulcerative colitis. Neuropsychologia, 1987, 25, 2: 367-374.
- Swaab DF, Chung WC, Kruijver FP, Hofman MA, Ishunina TA. Sexual differentiation of the human hypothalamus. Adv Exp Med Biol., 2002, 511:75-100.
- Swaab DF. Neuropeptides in hypothalamic neuronal disorders. Int Rev Cytol., 2004, 240: 305-75.
- Swaab DF. Sexual orientation and its basis in brain structure and function. Proc.Nat.Acad.USA, 2008, July 29; 105, 30: 10273-10274.
- Tugnet N, Goddard JC, Vickery RM, Khoosal D, Terry TR. Current management of male-to-female gender identity disorder in the UK. Postgrad Med J., 2007, Oct; 83, 984: 638-642.
- Weinstock M. Sex-dependent changes induced by prenatal stress in cortical and hippocampal morphology and behaviour in rats: an update. Stress, 2011, Nov; 14, 6: 604-13.
- Yokota Y, Kawamura Y, Kameya Y. Callosal shapes at the midsagittal plane: MRI differences of normal males, normal females, and GID. IEEE Engineering in Medicine and Biology, 2005, 27th Annual Conference, p.3055.

SkePSYs
Ana Giurgiuca
Din 16 până în 22 martie s-a sărbătorit „Săptămâna Creierului“, în Bucureşti fiind organizată o conferinţă, pentru publicul larg, cu subiectul: „Creierul ca fundament al creativităţii“. Vă propun să dezlegaţi jocurile de mai jos şi să aflaţi mai multe despre creativitate, acest atribut al umanităţii, a...
Dansul şi inteligenţa chinestezică
Emil-Daniel Alexe
Într-un trecut relativ apropiat, dansul şi psihoterapia s-au îmbinat fructuos în terapia prin dans şi miscare, o terapie folosită azi curativ la nivel emoţional şi mental. Având o experienţă personală în ceea ce priveşte dansul, m-am întrebat: Ce ajută un dansator să fie mai bun decât un alt dansator? Cum se...
Disfuncţiile comunicării în ADHD
Rareș Ignat
Obiective: O evaluare recentă a raportului dintre intervenţiile medicamentoase şi cele psihologice în ADHD relevă existenţa unei abordări reducţioniste în iniţierea terapiei medicamentoase. Obiectivul acestui articol este să evalueze care sunt principalele elemente ce au dus la dezechilibrul dintre intervenţ...
Depresia provine de la neanderthalieni?
Dan Pereţianu, Aurelian Nebel, Sorin Şuba, Denis Păduraru
Circa 2-4% din genomul uman conţine gene neanderthaliene. Aceasta sugerează că H. Neanderthaliensis s-a sublimat în H. sapines, şi că nu a dispărut acum 30 000 de ani. Inserţia de gene neanderthaliene la Homo sapiens a condus la exemplare care au fost mai adaptate regimului de temperaturi reci al Europei şi ...
Despre rost și depresie. Scurte consideraţii lingvistico-medicale
Dan Pereţianu, Aurelian Nebel, Sorin Şuba, Denis Păduraru
Depresivii afirmă deseori: „Viaţa nu mai are rost“. Acest înţeles al cuvîntului „rost“ nu există în alte limbi; i.e., engleza şi franceza, „rostul“ este „semnificaţie“, „sens“. În limba română, „rostul“ vieţii este altce...